【综述】中枢神经系统淋巴引流在缺血性卒中中的研究进展
时间:2026-05-05 17:07:54 热度:37.1℃ 作者:网络
摘要:缺血性卒中为高发病率、高致残率、高病死率的脑血管疾病。中枢神经系统淋巴引流在维持大脑内环境稳态、调控免疫功能等方面发挥重要作用,为缺血性卒中后的病理机制探索与临床治疗开辟了新视角。淋巴引流系统可通过胶质淋巴系统物质交换路径、脑膜淋巴管及颈深淋巴结引流通道协同调控脑脊液循环,清除代谢废物并转运免疫细胞,参与缺血性卒中后脑水肿、血-脑屏障破坏及神经炎症等关键病理环节。该文拟综述中枢神经系统淋巴引流系统的组成与功能及其在缺血性卒中发展过程中的作用机制,以期为缺血性卒中治疗提供新的研究方向。
缺血性卒中(ischemic stroke, IS)是由于脑动脉狭窄或闭塞造成局部脑血流量减少引发的脑组织急性缺血、坏死,是全球范围内造成死亡和残疾的重要原因之一[1],中枢神经系统(central nervous system, CNS)淋巴引流是调节脑内稳态和免疫功能的重要途径,在维持脑内液体平衡、抵抗 CNS疾病等方面具有重要作用,CNS淋巴引流系统包括胶质淋巴系统(glymphatic system, GS)、脑膜淋巴管(meningeal lymphatic vessels, mLVs)和颈深淋巴结(deep cervical lymph nodes, dCLNs),三者通过“废物清除-淋巴引流-免疫激活”的步骤协同运作[2-3],其功能异常与 IS等脑血管疾病的发展关系密切[4-5]。笔者拟综述 CNS淋巴引流系统的组成与功能及其在 IS发展过程中的作用机制,以期为 IS治疗提供新的研究方向。
1 CNS淋巴引流系统的组成与功能
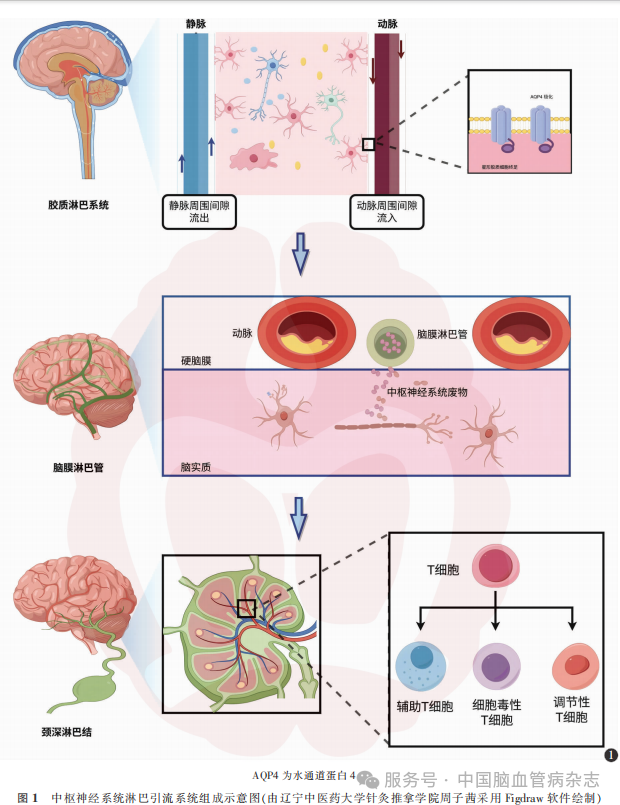
脑脊液主要由侧脑室脉络丛上皮细胞分泌[6],依赖钠钾泵离子梯度及碳酸氢盐协同向外转运,经脑室系统进入蛛网膜下腔后,大部分经蛛网膜颗粒吸收最终回流入血,剩余脑脊液则向脑实质深层灌注并参与 CNS代谢废物清除[7]。GS为存在于脑实质内的液体交换系统,负责驱动脑脊液与组织间质液进行物质交换;mLVs为存在于硬脑膜内的结构化引流网络,负责将脑内免疫细胞及脑源性代谢废物定向引流至 dCLNs;在 dCLNs中,免疫细胞通过抗原提呈作用激活免疫应答,代谢废物则经淋巴液进入外周淋巴循环系统被清除,实现 CNS免疫监视与代谢清除的功能闭环[8-9]。
1.1 GS:脑脊液-脑组织间质液交换通路
GS又称类淋巴系统,为 CNS中依赖星形胶质细胞终足表面水通道蛋白 4(aquaporin-4, AQP4)介导的脑脊液-间质液双向交换通路,Iliff等[10]首次在小鼠脑内观察到该液体运输路径。GS主要包括以下 3个关键阶段[10-11]:(1)脑脊液沿动脉周围间隙流入;(2)AQP4介导物质交换;(3)代谢废物沿静脉周围间隙流出。上述过程障碍可导致 β-淀粉样蛋白(β-amyloid protein, Aβ)、tau蛋白等产物清 除受阻,引发神经毒性物质在脑实质内异常积聚,因此 GS功能异常与神经系统疾病密切相关[12]。
1.1.1 脑脊 液 沿 动 脉 周 围 间 隙 流 入:2008年,Carare等[11]采用荧光示踪剂追踪技术验证了脑脊液的流动通道,其可通过软脑膜与星形胶质细胞终足覆盖的胶质界膜形成脑血管周围空间,并沿脑穿支动脉深入脑实质。作为脑膜与脑血管系统之间的流动通道,动脉周围间隙内液体运动受动脉搏动驱动,其流动峰值与心电 R波高度同步[13]。此外,呼吸周期、血管平滑肌细胞“微泵”作用及神经血管耦合等因素也协同参与调控脑脊液灌注过程,确保营养物质稳定输送及脑内环境动态平衡[14-15]。
1.1.2 AQP4介导物质交换:Iliff等[10]在其早期关于脑内液体循环通路研究的基础上,通过特异性敲除小鼠 AQP4基因以明确 Aβ的清除路径,结果显示,与野生型小鼠比较,特异性敲除 AQP4基因小鼠脑脊液示踪剂卵清蛋白耦联 Alexa Fluor 647(ovalbumin-conjugated Alexa Fluor 647, OVA-A647)向脑实质的转移量减少(P<0.05)且间质溶质清除率下降约 70%(P<0.01),提示 AQP4在 GS清除功能中发挥重要作用。靶向调控星形胶质细胞终足上的 AQP4可能是维持脑内液体稳态、促进废物清除的关键途径[16-18]。
星形胶质细胞为 CNS中数量最多的神经胶质细胞类型,其突起末端形成的终足结构是血-脑屏障的重要组成部分,AQP4为其终足上的选择性膜整合蛋白,可高效介导水分子跨膜转运[19-20]。该蛋白在脑内呈高度特异性分布,在星形胶质细胞终足朝向脑血管腔或脑实质区域尤为密集,在星形胶质细胞胞体或其他部位表达较低,这种不对称性定位分布称为极性分布,当 AQP4表达量降低或消失,并弥散到胞体及其他部位时,则可出现 AQP4极性丢失[21]。肌营养不良蛋白-糖蛋白复合物(dystrophin-glycoprotein complex, DGC)为连接细胞外基质与细胞内骨架的跨膜蛋白复合物,可特异性识别、结合 AQP4胞内结构域,为 AQP4持续富集于星形胶质细胞终足提供稳定锚定位点,这一特性称为锚定作用,当该复合物成分表达量发生变化或结构异常时可导致 AQP4极性丧失,影响其液体转运功能与 GS清除效率[22]。
除蛋白本身的结构基础外,多种信号通路可共同调控 AQP4的功能。白细胞介素(IL)1α、IL-1β及肿瘤坏死因子 α(TNF-α)等促炎因子可激活核因子κB(nuclear factor kappa-B, NF-κB)等转录因子,启动基质 金 属 蛋 白 酶 等 靶 基 因 蛋 白 表 达 并 破 坏DGC,导致 AQP4极性丧失和脑间质溶质清除率下降[23]。上述促炎因子亦可激活丝裂原活化蛋白激酶(mitogen - activatedproteinkinase,MAPK), 通过磷酸化修饰或影响该复合物结合力介导 AQP4极性表达,进而抑制 GS清除功能,引发脑能量代谢紊乱和神经元损伤[24]。
1.1.3 代谢废物沿静脉周围间隙流出:静脉周围间隙是软脑膜连接脑小静脉穿出脑实质时形成的潜在空隙,与动脉周围间隙一起构成 GS液体引流通道[10]。脑脊液完成对流交换后形成的脑脊液-间质液混合液在动脉搏动、静脉回流、呼吸负压及 AQP4功能等作用下可沿静脉周围间隙定向流动并排出;而睡眠-觉醒周期对 GS清除效率具有调节作用,睡眠状态下,脑血容量周期性波动及血管周围间隙扩张可驱动脑脊液流量增加,促进脑脊液与间质液交换,从而提高 GS的液体流速及 Aβ等神经毒性蛋白的清除效率[25-26]。
1.2 mLVs:淋巴引流网络
mLVs是硬脑膜内具有典型淋巴管结构特征的管状网络,其内皮细胞可表达 Prospero同源异形盒蛋白1(prospero-related homeobox prorein 1, PROX1)、血管内皮生长因子受体 3(vascular endothelial growth factor receptor 3, VEGFR3)等 淋 巴 内 皮 细 胞 标 志物[27],2015年,Aspelund等[9]在 PROX1-GFP转基因小鼠及 VEGFR3信号缺陷小鼠模型中观察到硬脑膜内存在淋巴管结构且可直接引流脑内大分子物质,同年 Louveau等[28]小鼠实验进一步证实,mLVs可介导 T细胞等脑内免疫细胞向外周免疫系统转运,打破了“CNS无淋巴管”的传统认知,填补了脑内废物如何引流至外周循环的研究空白。
1.2.1 脑脊液与代谢废物引流:2017年,Absinta等[29]通过高分辨率 MRI在人体中观察到 mLVs,其主要位于颅内硬脑膜层,常与脑膜中动脉、静脉及颅神经伴行分布,mLVs内皮细胞以独特的“纽扣状”形式进行连接,以防止液体反流,其为脑脊液摄取及脑内代谢废物定向外排的关键通道。PROX1作为 mLVs内皮细胞的关键转录调控因子,是推动和鉴别血管内皮细胞向 mLVs内皮细胞分化的特异性分子标志物,而 mLVs结构成熟及引流功能的实现还需依赖血管内皮生长因子 C(vascular endothelial growth factor C, VEGF-C)与 其 受 体 VEGFR3的 持 续 作用[30]。Antila等[31]通过敲除小鼠 VEGF-C基因或抑制 VEGF-C/VEGFR3通路进一步证实,阻断该通路可导致 mLVs发育不良、退化甚至缺失。
1.2.2 免疫监视启动与调控:除了转运脑内液体及代谢废物外,mLVs还可引导免疫细胞与抗原定向迁移,在生理状态下,mLVs持续将脑脊液及其所含有的低浓度脑源性抗原引流至 dCLNs,使持续性、低水平的抗原信号传递至外周淋巴系统,这种稳态下的抗原提呈有助于维持 CNS的免疫耐受,从而降低异常免疫激活及自身免疫疾病发生风险[28]。病理状态下,树突状细胞等抗原提呈细胞可通过胞吞作用摄取并加工抗原,随后依赖 mLVs转运至外周[32]。mLVs内皮细胞通过分泌趋化因子配体 21与免疫细胞表面趋化因子受体 7结合等形式,引导树突状细胞及初始 T细胞定向迁移至 dCLNs,激活适应性免疫应答[33]。此外,mLVs还可通过影响 T细胞激活强度和脑内炎症微环境间接调控效应细胞向损伤区域的再迁移效率[34]。
1.3 dCLNs:免疫应答枢纽
dCLNs位于胸锁乳突肌深部,沿颈内静脉分布,是 CNS抗原、免疫细胞及代谢废物的外周中转站,来自 mLVs的淋巴液经输入淋巴管流入被膜下淋巴窦,依次流经皮质窦、髓窦,最终经输出淋巴管汇入颈静脉淋巴干回流至体循环;dCLNs滤过及免疫功能可拦截或清除 CNS异常抗原等有害成分,避免其入血扩散,起到维系机体稳态、减轻 CNS损伤的关键作用[35]。
1.3.1 淋巴液滤过与清除:Da Mesquita等[36]研究显示,Aβ等大分子代 谢 废 物 可 经 mLVs引 流 至dCLNs,dCLNs淋巴窦中的条索结构可增大与淋巴液接触面积并减缓其流速,使巨噬细胞更好地捕捉和清除 Aβ。研究显示,结扎阿尔茨海默病小鼠 dCLNs后,其大脑皮质及海马区均出现大量 Aβ沉积和AQP4极性丧失,并伴随着小鼠认知缺陷程度和探索行为受限加重[37]。上述研究表明,dCLNs是 mLVs的核心引流节点,其结构完整和功能完备对脑源性代谢废物的有效清除和脑内环境稳态调控具有重要意义。
1.3.2 抗原识别与免疫应答:除执行基础淋巴液滤过功能外,dCLNs亦是外周免疫识别和应答的关键场所。脑源性抗原可经 mLVs引流至 dCLNs,树突状细胞将其捕获、处理并呈递给 T细胞,激活的效应 T细胞在趋化因子及黏附分子介导下经血液循环穿过血-脑屏障回巢至 CNS病灶[34]。效应 T细胞可分化为辅助 T细胞、细胞毒性 T细胞和调节性T细胞,辅助 T细胞可分泌干扰素 γ、TNF-α等细胞因子调控炎症,细胞毒性 T细胞可释放穿孔素和颗粒酶介导靶细胞杀伤,调节性 T细胞可分泌转化生长因子 β等成分抑制自身免疫反应,维持免疫耐受[38]。目前研究认为,CNS与外周淋巴组织之间的B细胞群体存在动态交流[39-40]。Eggers等[39]通过流式细胞术及免疫组库测序等技术在多发性硬化症患者脑脊液和外周血中证实了 CNS与外周 B细胞群体间存在克隆关系。Stern等[40]观察到脑膜、脑脊液及脑实质中 的 B细 胞 与 dCLNs同 源,提 示dCLNs可能是脑内致病性 B细胞完成克隆扩增、亲和性成熟并返回 CNS的关键免疫场所。
CNS淋巴引流系统的组成示意图见图 1。
2 IS后调控 CNS淋巴引流系统的作用
氧化应激、炎性反应是 IS进程中引发神经元死亡与脑损伤的主要病理损伤机制,IS后血-脑屏障破坏与脑水肿形成进一步加剧神经功能衰退及认知障碍[41]。研究表明,IS后通过介导 GS功能、改善mLVs-dCLNs引流来调控 CNS淋巴引流系统,清除梗死组织及代谢毒物,进而减轻 IS后神经损伤,促进组织修复[42-44]。
2.1 双向调控脑水肿
脑水肿是脑内液体异常蓄积所致脑组织肿胀的病理状态,根据血-脑屏障受损程度及病理特征可将其分为细胞毒性水肿与血管源性水肿;CNS淋巴引流系统可通过调节水分跨膜转运、外周引流等环节对脑水肿的发生发展产生阶段性影响[45]。
细胞毒性水肿发生在缺血数分钟至数小时内,缺血早期能量代谢障碍可导致钠钾泵功能失常,细胞内 Na+潴留,K+外流,触发去极化,引发神经元及星形胶质细胞肿胀[45]。Mestre等[46]构建小鼠大脑中动脉闭塞模型,通过静脉和脑池注射22Na及3H-甘露醇以判断水肿情况及水肿液来源,并基于 MRI确定脑脊液浸入组织位置,结果显示,造模前小鼠双侧脑半球 示 踪 剂 流 入 量 差 异 无 统 计 学 意 义 (P>0.05),而造模后缺血侧示踪剂流入量较对侧增加约3倍(P<0.01),并主要沿 GS中脑穿支动脉流入(97.7%;沿脑小静脉流入仅占 2.3%,P<0.01)经血管周围间隙浸润脑实质;静脉注射22Na及3H-甘露醇后其在缺血侧与对侧的积聚量差异无统计学意义(均 P>0.05),但经脑池注射后,缺血侧22Na(P=0.013)及3H-甘露醇(P=0.014)积聚量均较对侧增加,提示脑脊液沿 GS异常流进可能是 IS早期脑水肿的诱因。Lyu等[47]构建大脑中动脉闭塞再灌注大鼠模型,并进行免疫荧光、免疫组化、脑含水量检测,结果显示,与假手术组相比,模型组梗死侧皮质及海马区 AQP4极化分布紊乱,密度增加(P<0.01),星形胶质细胞终足肿胀,脑含水量增加(P<0.01),提示 AQP4极性紊乱及跨膜水转运增强参与细胞肿胀过程,但其作用可能与后续血管源性水肿相关。
血管源性水肿通常发生在梗死后数小时至数天内,血-脑屏障受损使血管通透性增加,血浆蛋白、离子和水分外渗至脑外间隙,在此过程中小胶质细胞激活及星形胶质细胞增殖进一步加剧了 AQP4极性丧失与脑水肿程度[48]。Yang等[49]构建小鼠大脑中动脉闭塞再灌注模型,于再灌注1、24h及3、7d进行取材,通过免疫印迹、免疫荧光检测 AQP4定位、反应性星形胶质细胞及脑含水量变化,并追踪脑脊液示踪剂荧光强度变化以评估 CNS淋巴引流能力,采用单因素方差分析及独立样本 t检验计算组间差异,效应量以平均差(mean deviation, MD)及 95% CI表示,结果显示,与假手术组相比,模型组脑含水量于再灌注 1h及 3d出现高峰,其中再灌注 3d时脑含水量明显增加 (MD=0.177,95% CI:0.123~0.232,P< 0.01)、脑内示踪剂残留面积增大(MD=17.073,95% CI:8.211~25.935,P= 0.003);该研究将再灌注 3d界定为血管源性水肿阶段,并分析AQP4极性及 DGC中核心成分肌营养不良蛋白 71的变 化 情 况,结 果 显 示,在 该 阶 段,模 型 组 小 鼠dCLNs示踪剂荧光强度较假手术组降低(MD=-0.506,95% CI:-0.618 ~ 0.395, P<0.01),AQP4极化丧失呈弥散分布,反应性星形胶质细胞内胶质纤维酸性蛋白相对密度增加(MD=0.507,95% CI:0.085~0.930,P= 0.013),提示反应性星形胶质细胞增殖;通过病毒上调肌营养不良蛋白71,可促进 AQP4极性恢复,增强 dCLNs示踪剂荧光强度(MD=0.245, 95% CI:0.004~0.485, P=0.048),减少脑含水量(MD=-0.127,95%CI:-0.243 ~ 0.011,P=0.027)。上述研究结果提示,AQP4极性丧失可能加重血管源性水肿阶段的液体潴留,维持 DGC结构及稳定表达有助于改善脑内液体动力学失衡。
2.2 修复血-脑屏障
血-脑屏障由脑微血管内皮细胞与基底膜、周细胞及星形胶质细胞足突等成分构成,是维持脑内微环境稳态的重要结构基础[50]。Müller等[51]纳入138例卒中患者(IS103例,出血性卒中 35例)进行回顾性分析,结合住院期间 CT影像学分析水肿变化、脑脊液与血浆白蛋白比值以评估血-脑屏障功能,结果显示,与无血-脑屏障受损患者(90例)相比,血-脑屏障受损患者(48例)调整后相对水肿体积(脑水肿体积/梗死体积)增加[4.0(95% CI:2.8 ~ 4.5)比 2.3(95%CI:1.8~3.0),P=0.037],神经功能恢复(发病后 3个月内改良 Rankin量表评分较出院时降低至少 1分)率降低[41.7%(20/48)比56.7%(51/90);aOR=0.37,95% CI:0.16 ~ 0.88, P=0.025],提示血-脑屏障损伤程度与卒中预后密切相关。
IS发生后,缺血缺氧环境诱导活性氧及 IL-1β等炎症因子大量释放,促使内皮细胞间紧密连接蛋白降解,基质金属蛋白酶等对基底膜产生持续损伤,血管结构稳定性降低,导致神经毒性物质沉积,从而加重 IS后脑损伤[52]。星形胶质细胞足突结构亦在该过程中遭到破坏,导致 DGC异常、AQP4极性丧失,降低 GS清除效率,影响脑内代谢废物经 mLVs-dCLNs通路的后续引流[53]。Capuk等[54]构建星形胶质细胞特异性敲除 NBCe1基因小鼠,并对其进行短暂性大脑中动脉闭塞处理,通过 2,3,5-三苯基氯化四唑染色、MRI及 T2加权成像评估脑梗死体积与水肿情况,采用免疫荧光检测 AQP4极性,采用伊文思蓝染 色 检 测 血-脑 屏 障 通 透 性,结 果 显 示,与NBCe1基因敲除 IS小鼠比较,野生型 IS小鼠脑梗死体积较大[(60.8±19.1)%比(13.6±9.1)%,P<0.05],脑水肿加剧[半球肿胀程度:(同侧半球体积-对侧半球体积)/对侧半球体积;(16.2±4.6)% 比(4.8±2.0)%,P<0.05],脑损伤区 NBCe1表达上调[NBCe1簇计数:(163.8±45.2)个比(40.5±9.2)个, P= 0.01],AQP4极性丧失并弥散分布至实质区域,血-脑屏障通透性增加[(262±23)个细胞/视野比(123±46)个细胞/视野,P< 0.05],与其相比,NBCe1基因敲除 IS小鼠脑组织中 AQP4呈极性分布,血-脑屏障通透性降低[(123±46)个细胞/视野比(262±23)个细胞/视野,P<0.05],基质金属蛋白酶抑制剂表达上调(1.53±0.67)倍(P<0.01)。上述研究结果提示,AQP4极性分布在血-脑屏障保护中具有重要作用,通过靶向调控 AQP4极性、恢复星形胶质细胞功能,或将有利于促进 AQP4介导的 GS发挥液体交换与代谢废物清除功能。研究显示,IS小鼠模型脑损伤区细胞外液对皮质和海马原代神经元的毒性作用可持续至少 7周,提示卒中后毒性废物沉积可导致组织长期损伤,影响血-脑屏障结构稳定及神经功能恢复[55]。Kim等[56]构建大脑中动脉闭塞再灌注小鼠模型,探究混合电光刺激干预后小鼠 GS功能及脑损伤情况,通过水迷宫实验评估其学习能力,采用脑脊液示踪剂检测 GS功能,采用 MRI、免疫荧光及免疫印迹检测 AQP4及 IL-1β表达,结果显示,与对照组相比,IS组小鼠寻找平台时间延长(P<0.05),脑脊液示踪剂浸润脑实质量降低,AQP4表达升高、极化降低(均 P<0.01);与 IS组相比,混合电光刺激组小鼠寻找平台时间缩短,脑梗死体积减小,脑脊液示踪剂流入脑实质量升高,缺血皮质神经元数量增加,AQP4极性表达升高,IL-1β表达降低(均 P<0.05),上述研究结果提示,通过调节AQP4极性分布或可改善 GS功能,促进 IS后神经功能恢复,降低 IL-1β等炎症因子水平,减轻血-脑屏障损伤。
2.3 动态调节神经炎症
神经炎症贯穿 IS发展的全过程,缺血区域坏死细胞可释放损伤相关分子模式激活小胶质细胞及星形胶质细胞,并促进中性粒细胞、淋巴细胞等募集及TNF-α、IL-1β等促炎因子大量释放[57]。CNS淋巴引流系统与神经炎症间可能存在动态调控关系,其作用呈阶段性变化。
IS急性期(通常指发病 72h内),损伤相关分子模式通过激活 NOD样受体热蛋白结构域相关蛋白3等方式促进小胶质细胞向促炎表型 M1型极化,释放 IL-1β、IL-18等促炎因子,脑缺血-再灌注损伤发生后,免疫细胞可通过血-脑屏障进入脑实质,进一步加重炎症反应[57]。Jiang等[58]纳入 34例首发 IS患者并检测其与对照组(30例健康志愿者)外周血中长链非编码核糖核酸生长停滞特异性转录本 5 (long non-coding ribonucleic acid growth arrest-specific 5, lncRNA GAS5)、信 号 转 导 与 转 录 激 活 因 子 5A(signal transducer and activator of transcription 5A, STAT5A)表达,实时荧光定量聚合酶链式反应、免疫印迹结果显示,与对照组比较,IS组上述指标均升高(均 P<0.01);该研究构建脑缺血-再灌注大鼠模型及星 形 胶 质 细 胞 氧-葡 萄 糖 剥 夺/复 氧 模 型 探 究STAT5A/AQP4通路调控炎症相关的机制,通过2,3,5-三苯基氯化四唑染色、脑含水量检测其脑梗死体积及水肿情况,结合实时荧光定量聚合酶链式反应、免疫印迹检测 GAS5、STAT5A、AQP4及促炎细胞因子 IL-1β、IL-6、TNF-α表达,CCK-8法检测细胞活力,结果显示,与对照组比较,模型组大鼠脑梗死体积增大,脑含水量增加 (均 P<0.01),GAS5、STAT5A及 AQP4表达均上调(均 P<0.01),IL-1β、IL-6及 TNF-α表达均增加 (均 P<0.01);抑 制STAT5A可降低脑缺血-再灌注大鼠脑梗死体积、脑含水量及 AQP4表达(均 P<0.01),敲低 AQP4可提高星形胶质细胞的细胞活力,降低 IL-1β、IL-6及TNF-α表达(均 P<0.01)。上述研究表明,通过STAT5A轴抑制 AQP4表达,可能有利于降低 IS后神经元凋亡和促炎因子水平,缓解组织损伤。Lee等[59]在脑缺血-再灌注小鼠模型中破坏 mLVs,免疫荧光及流式细胞术分析显示,与卒中小鼠相比,mLVs受损的模型小鼠脑内 CD4+、CD8+T细胞、中性粒细胞及单核细胞向缺血脑组织的浸润减少 (均 P<0.05),脑梗死体积减小(P<0.05),通过单细胞RNA测序技术观察结果显示,与卒中小鼠比较,mLVs破坏可特异性降低 dCLNs淋巴内皮细胞富含半胱氨酸血管诱导因子 61表达(P<0.01),该结果提示 mLVs-dCLNs可能通过调控富含半胱氨酸血管诱导因子 61表达影响脑内炎症反应进程。
随着病程进入亚急性期恢复阶段(IS后数周至数月),部分小胶质细胞向抗炎表型 M2型转化,转化生长因子 β、IL-10等抗炎因子表达增加,并通过修复受损细胞、促进吞噬作用等方式参与神经修复及免疫调节[57]。一项影像学研究采用沿血管周围间隙的扩散张量成像分析 (diffusion tensor image analysis along the perivascular space, DTI-ALPS)检测IS患者(51例)康复训练前及康复训练后 30d的DTI-ALPS指数以评估 GS功能,结果显示,与康复训练前相比,IS组患者康复训练 30d后梗死侧 DTI-ALPS指数增加(1.09±0.22比 0.98±0.30,P<0.05),提示血管周围液体交换功能部分恢复,GS功能障碍缓解[60]。研究表明,IS过程中产生的炎症介质可持续作用于星形胶质细胞,使其发生反应性增殖、肥大,并迁移至损伤周围聚集形成胶质瘢痕,急性期胶质瘢痕的形成有助于限制炎症细胞向脑组织浸润,降低炎症范围,但其在亚急性期及恢复阶段将阻碍神经轴突再生与组织重塑[61]。在上述过程中,调控 AQP4极性表达或可影响星形胶质细胞在脑损伤区域增殖和聚集,进而介导胶质瘢痕形成的范围、密度,直接影响神经炎症微环境稳态及神经功能修复进程[62]。Tian等[63]构建短暂性大脑中动脉闭塞大鼠模型,于建模后 1、3、7、14、28d通过 MRI、免疫荧光检测探究 DTI-ALPS指数变化及 AQP4极化情况,结果显示,与对照组相比,模型组大鼠双侧 DTI-ALPS指数于 IS急性期(造模后第 1天)降低(P<0.01),随后该指标于造模后第 3、7天略有回升,但差异均无统计学意义(均 P>0.05);模型组大鼠缺血侧建模后第 14天 DTI-ALPS指数均高于建模后第1、3、7天(均P<0.01),建模后第 28天 DTI-ALPS指数均高于建模后第 1、3、7、14天(均 P<0.05);对建模后第 28天脑梗死周围区域进行免疫荧光检测,结果显示,与对照组相比,模型组大鼠梗死核心区附近星形胶质细胞出现明显增殖、迁移,形成胶质瘢痕并伴随 AQP4极性分布紊乱,胶质瘢痕周围区域可见AQP4极性分布丧失且 AQP4极性与 DTI-ALPS指数随时间的变化趋势基本一致,均于建模后第 14、28天较早期时间点增加[与建模后第 1、3、7天比较,建模后第 14天 AQP4极性均增加;与建模后第 1、3天比较,建 模 后 第 28天 AQP4极 性 均 增 加 (均 P<0.01)。上述结果提示,于 IS亚急性期恢复阶段介导 AQP4极性恢复,可能有利于改善 GS功能,进而影响胶质瘢痕形成及周围区域损伤修复。Yanev等[64]通过 VEGFR3突变诱导小鼠 mLVs发育缺陷,并构建短暂性大脑中动脉闭塞模型,结果显示,与野生型IS小鼠相比,VEGFR3突变诱导 mLVs发育缺陷模型小鼠 mLVs密度降低(P<0.01),脑梗死体积增大[(24.3±12.2)mm3比(9.7±6.9)mm3,P<0.05],提示 mLVs缺陷可能进一步加剧 IS后组织损伤,支持了mLVs结构完整及功能完备在 IS预后中的关键作用。
2.4 引导与启动免疫应答
IS后,脑损伤区特异性抗原及炎症介质释放增加,血-脑屏障完整性破坏,巨噬细胞、中性粒细胞等免疫细胞迅速聚集于脑组织损伤区域,启动免疫应答,引起中枢及外周免疫微环境改变[65]。CNS淋巴引流系统在这一过程中不仅参与抗原清除,还发挥抗原转运与免疫启动的关键功能。
研究表明,T细胞等免疫细胞可通过血-脑屏障进入血管周围间隙,损伤组织释放的抗原及炎症介质也可经过 GS物质交换途径流向脑膜及蛛网膜下腔[66]。部分树突状细胞摄取抗原后,亦可沿 mLVs向 dCLNs迁移,dCLNs中激活的免疫细胞释放细胞因子并经 mLVs反向运回脑内,参与局部免疫调节过程[66-67]。Bai等[68]在大脑中动脉闭塞大鼠模型建模后 14d通过追踪示踪剂 OVA-A647引流至 mLVs与 dCLNs含量评估淋巴引流功能,结合免疫荧光检测评估 T细胞浸润及神经元损伤情况,结果显示,与健康大鼠相比,模型组大鼠 mLVs与 dCLNs中示踪剂 OVA-A647含量均下降(均 P<0.01),提示淋巴引流通路受阻,缺血区 CD4+、CD8+T细胞浸润均增加(均 P<0.01),并伴随神经元丢失、损伤程度加剧;通过颅骨转运手段对模型大鼠进行干预后,大鼠mLVs与 dCLNs中示踪剂 OVA-A647含量均升高(均 P<0.01),提示淋巴引流功能部分恢复,脑组织中 T细胞浸润均减少(均 P<0.05),脑室下区和齿状回神经干细胞数量均增加(均 P<0.01),提示恢复 mLVs-dCLNs引流功能有利于改善脑膜血供与局部微环境,影响 IS后 T细胞转运效率及相关免疫反应进程。此外,有研究通过切除双侧 dCLNs或脑室注射腺相关病毒-血管内皮生长因子 C增强淋巴引流,其在大脑中动脉闭塞大鼠模型中观察到了不同的结果,该研究采用 2,3,5-三苯基氯化四唑染色、MRI检测梗死体积,通过酶联免疫吸附试验、免疫组化观察 T细胞浸润情况,结果显示,与 IS组相比,切除 dCLNs卒中组大鼠脑梗死体积减小(P=0.0013),梗死侧 CD8+T细胞浸润在术后 3、12、72h均减少(P值分别为 0.0920、0.0025、0.0128),脾脏中CD4+、CD8+T细胞数量均于术后 3h降低(P值分别为0.0113、0.0002);与 IS组相比,脑室注射腺相关病毒-血管内皮生长因子 CIS组小鼠脑梗死体积扩大(P<0.01),梗死侧 CD4+、CD8+T细胞浸润差异均无统计学意义(均 P>0.05),脾脏中 CD4+、CD8+T细胞数量均于术后 3h增加(P值分别为 0.0280、0.0184)[69]。上述研究结果在梗死体积变化及免疫细胞浸润方面存在差异,提示淋巴引流机制在 IS后免疫应答阶段介导的促炎与免疫调节间的动态平衡机制仍不明确,精准调控淋巴引流功能或可成为干预 IS后免疫失衡的潜在策略。
3 总结
脑脊液在 GS驱动下与脑间质液对流交换,代谢废物、炎症介质及脑源性抗原经由 mLVs定向引流至 dCLNs,CNS淋巴引流系统在 IS的病理进程与稳态重建中至关重要。目前研究已初步明确增强该系统功能有助于加速 IS后水肿消除、降低毒性物质沉积并改善神经功能预后,然而其在 IS后不同阶段中的具体调控机制及如何将上述干预措施有效转化为临床治疗方案仍有待深入研究。未来可通过开展多中心、大样本的随机对照试验,纵向评估其对 IS后脑水肿消退、早期神经炎症标志物及长期神经功能恢复的影响,进一步明确其治疗时机,为减轻 IS后脑损伤、促进神经功能修复提供新的治疗方向,推动IS综合治疗体系的发展与完善。

