慢性淋巴细胞白血病中治疗驱动的克隆演化及耐药机制
时间:2026-05-05 17:05:59 热度:37.1℃ 作者:网络
慢性淋巴细胞白血病(CLL)是一种以瘤内多样性和动态演化过程为特征的典型恶性肿瘤。高分辨率基因组分析表明,CLL的进展很少遵循线性轨迹;与之相反,其特点为由内在生物学特征和外部因素(包括治疗压力)塑造的复杂且不断演变的(亚)克隆结构。反复出现的基因改变影响关键信号通路和细胞过程,包括B细胞受体和NF-κB信号通路、DNA损伤反应、RNA加工和细胞凋亡。其中许多病变作为亚克隆事件出现并随后扩增,从而影响疾病进展、治疗耐药和转化。在过去十年中,CLL的治疗模式已从化学免疫治疗转向靶向药物,带来了显著的临床获益,但治疗耐药的出现仍然是一个重大挑战。
《Seminars in Cancer Biology》近日发表综述,总结了目前关于CLL克隆演化和耐药机制的知识。对化学免疫治疗的耐药通常由基因病变(如TP53畸变)和耐药微克隆的扩增驱动;而靶向治疗则选择不同的耐药机制,例如在接受BTK抑制剂治疗的患者中出现BTK和PLCG2突变,以及替代生存通路的激活。作者进一步讨论了新兴技术,包括单细胞测序和整合性多组学方法。最后,未来研究需要解决在不断变化的临床背景下(如联合靶向治疗、双特异性抗体和CAR-T细胞治疗)出现的耐药问题。综上所述,对克隆演化的深入理解对于制定个体化治疗策略和改善CLL患者的长期预后至关重要。

1.引言
多年来,化学免疫治疗(CIT),特别是氟达拉滨、环磷酰胺和利妥昔单抗(FCR)方案,一直是慢性淋巴细胞白血病(CLL)患者的标准治疗方案。然而在过去十年中,靶向治疗彻底改变了疾病管理模式。布鲁顿酪氨酸激酶(BTK)抑制剂(BTKi)伊布替尼是首个此类药物,可阻断BTK下游的B细胞受体(BcR)信号传导。另一项重大进展是BCL2抑制剂(BCL2i)维奈克拉的开发,它可恢复CLL细胞的凋亡。这两种药物最初被批准用于伴有TP53畸变的患者,现在已被广泛使用,并显示出显著的临床获益,特别是在具有未突变免疫球蛋白重链可变区(IGHV)基因等高危亚组中。
二代测序(NGS)技术揭示了CLL复杂的基因组图谱,识别出与侵袭性疾病、早期进展和较差生存相关的反复突变基因。克隆结构研究表明,虽然少数病变是早期驱动事件,但大多数以亚克隆形式出现,促进疾病演化和治疗耐药。NGS还加深了对CIT耐药的理解,揭示了关键通路和细胞过程中突变的富集。
尽管靶向药物疗效显著,但治疗耐药仍然是一个重大的临床挑战。在接受共价BTK抑制剂(cBTKi)治疗的患者中,耐药通常通过BTK中的点突变(阻止共价药物结合)或PLCG2中的突变(激活下游信号传导)发生。然而这些改变仅能解释约70%的复发案例,表明存在其他尚不明确的机制。对维奈克拉的耐药同样具有异质性:降低药物亲和力的BCL2 Gly101Val突变仅占难治性病例的一部分,提示存在其他分子逃逸路径。
在这篇综述中,作者总结了当前关于CLL克隆演化的知识,并探讨了驱动对传统CIT和现代靶向治疗产生耐药的分子机制。还讨论了对耐药生物学的理解如何为下一代BTK和BCL2抑制剂的开发提供信息。最后,本文提供了识别遗传耐药性以优化治疗策略和改善患者预后的临床相关性见解。
1.1. CLL中的克隆和亚克隆结构及演化
CLL的特征为成熟B淋巴细胞的克隆扩增,由细胞内在的遗传改变驱动,并由与肿瘤微环境(TME)的细胞外在相互作用维持。重要的是,CLL的进展很少是线性的,而是表现出复杂且动态的克隆结构,其中多个遗传上不同的亚克隆可以共存、竞争并随时间演化。这种瘤内异质性是疾病行为、进展和治疗耐药出现的基础。
1.1.1. 染色体改变作为克隆演化的早期驱动因素
对CLL克隆多样性的早期研究源于传统的核型分析和荧光原位杂交(FISH)分析,揭示了复发性染色体拷贝数改变(CNA)。11q和17p染色体区域的频繁缺失以及12号染色体三体与更具侵袭性的临床行为相关。相比之下,影响13q染色体的缺失在超过一半的患者中被发现,通常与更有利的预后相关。13q缺失和12号染色体三体通常在疾病起始时出现并作为创始事件持续存在,而高危病变(包括11q(ATM)和17p(TP53)缺失)则作为亚克隆事件在后面出现。在选择压力下(例如治疗期间和治疗后),这些亚克隆可以扩增并成为主导,导致更具侵袭性的临床行为、治疗耐药和更差的结果。纵向细胞遗传学研究还表明,染色体不稳定性促进持续的多样化,使同一患者体内多个亚克隆能够平行演化。
复杂核型(CK)通常定义为存在≥3种畸变,反映了演化中亚克隆累积获得的染色体增益、丢失和结构重排,通常与DNA损伤反应通路缺陷相关。CK可能在诊断时不存在,而在疾病进展过程中出现或从微小亚克隆扩增而来,与侵袭性生物学行为相关,包括更短的至首次治疗时间(TTFT)、对CIT的难治性、对靶向药物(至少在复发情况下)的反应较差,以及即使在无TP53异常的情况下也降低总生存期(OS)。与此概念一致,Baliakas及其同事证明,只有高核型复杂性(定义为存在≥5种畸变)才保留独立的负面预后意义。这些发现强调,CK不是一个单一的驱动病变,而是持续克隆多样化的复合标志物,反映了疾病的演化适应性和与不良临床结局的关联。
总的来说,这些观察结果确立了染色体异常不仅是静态的预后标志物,也是克隆演化的动态指标。因此,对染色体改变的FISH评估应在治疗开始前和任何后续治疗线时进行,而非在诊断时。
1.1.2. 芯片和NGS揭示的亚克隆结构
随着高分辨率单核苷酸多态性(SNP)芯片的引入,研究不仅证实了细胞遗传学发现,还识别了额外的CNA,包括2p或8q的增益以及9p或18p的缺失,所有这些都与不良结局相关。在2013年的一项里程碑式研究中,Landau及其同事将高密度SNP芯片与配对肿瘤-正常样本的全外显子测序(WES)相结合,系统地定义了CLL中的(亚)克隆结构;通过整合CNA与体细胞点突变,他们证明大多数CLL病例在诊断时已 harbors 多个遗传上不同的亚克隆[9]。驱动突变被证明可以作为早期克隆事件(例如12号染色体三体和13q缺失)或较晚的亚克隆事件发生,亚克隆驱动突变通常涉及关键基因,如TP53、SF3B1和ATM。重要的是,亚克隆驱动突变的存在和扩增与疾病进展和较差结局相关。对一部分病例的纵向采样进一步揭示,治疗施加了强大的选择压力,导致亚克隆组成随时间动态变化,并将亚克隆复杂性确立为CLL的核心特征。
1.1.3. 复发性突变信号通路中的克隆演化
NGS技术的应用迅速揭示了CLL的基因组图谱,表明复发突变影响多个信号通路和过程。最常改变的通路包括:
-
BCR和NF-κB信号通路(例如EGR2、NFKBIE、MYD88):通过微环境相互作用增强存活和增殖;
-
DNA损伤反应和细胞周期控制(例如TP53、ATM、POT1):其破坏导致基因组不稳定性;
-
RNA加工和核糖体功能(例如SF3B1、XPO1、RPS15):影响转录保真度和细胞应激反应;
-
NOTCH和MAPK信号通路(例如NOTCH1、BRAF):促进生长和免疫逃逸。
在诊断时,CLL通常表现为一个主导的创始克隆,伴随携带额外基因突变的较小亚克隆。这些亚克隆可能保持稳定、扩增或减少,取决于选择压力,尤其是治疗。纵向基因组研究表明,虽然某些驱动突变(如NOTCH1、SF3B1、NFKBIE、RPS15、BIRC3和TP53中的突变)在诊断时可能以低变异等位基因频率(VAF)存在,但它们通常在疾病进展或治疗压力下扩增,从而影响疾病结局。
然而,这些改变的生物学和临床影响因基因而异。例如,TP53破坏(通过突变或17p缺失)会损害细胞凋亡,并与侵袭性疾病、对CIT的难治性和较差生存密切相关。NOTCH1突变促进白血病细胞存活,并与对抗CD20单克隆抗体的耐药相关,而参与RNA剪接的SF3B1突变与更快的进展和不良预后相关。类似地,BIRC3和ATM的突变也导致治疗耐药。BIRC3突变在诊断时通常是亚克隆的,但可以随时间扩增,经常与ATM改变同时发生。携带BIRC3突变的患者TTFT更短,CIT后的无进展生存期(PFS)更差。NFKBIE突变通常在疾病进展过程中作为亚克隆事件出现。通过破坏NF-κB信号通路的抑制性控制,这些突变增强了促存活和微环境依赖的信号,使得突变亚克隆即使在没有治疗的情况下也能逐渐扩增。EGR2突变通常是早期的克隆性或近克隆性事件;在功能上,它们影响与BCR信号和分化相关的转录程序,在IGHV未突变的CLL(U-CLL)中富集,并与侵袭性疾病和不良预后相关。相比之下,RPS15突变代表较晚的演化事件,在诊断时罕见,但在复发时(尤其是在CIT后)经常检测到;它们影响核糖体功能并损害p53介导的应激反应,赋予对基因毒性损伤的抵抗力,并使治疗难治性克隆能够击败敏感群体。
重要的是,多个信号通路通常在同一患者体内但不同亚克隆中被改变,反映了分支演化和通路冗余。这使得白血病细胞能够适应选择压力,如免疫监视和治疗。最近的研究还表明,单个CLL克隆内驱动突变所改变的不同信号通路的数量本身具有预后意义:其白血病细胞 携带影响多个通路的突变的患者表现出更短的TTFT、增加的进展风险和较差的生存,且独立于个别高危基因。
2. 微克隆作为耐药的储存库
微克隆,即低于传统Sanger测序检测限的低频率白血病细胞亚克隆群体,已被证明在CLL中高度相关。携带TP53突变的白血病细胞微克隆已被证明影响对CIT(尤其是FCR方案)的反应。即使以非常低的VAF存在,TP53畸变也会赋予对这些药物的耐药性,导致与缺乏TP53缺陷的患者相比,PFS更短,OS更差。使用灵敏方法(如深度测序或数字PCR)的研究表明,这些微克隆通常随时间或在治疗压力下扩增,最终驱动复发。目前尚不清楚携带TP53突变的微克隆在靶向治疗后如何受到影响,因为这些治疗施加了与CIT不同的选择压力。对于携带SF3B1、ATM、BIRC3改变的微克隆也有类似的报道。因此,尽管在诊断时通常检测不到,但这些微克隆充当了瘤内异质性的储存库,可以选择性地随时间或在治疗压力下扩增,支撑着适应性演化、治疗失败、疾病进展和转化。
2.1. 表观遗传学与克隆演化
表观遗传改变通过在不改变DNA序列的情况下塑造细胞身份、可塑性和适应性,在CLL的克隆演化中发挥核心作用。全基因组DNA甲基化和染色质分析研究表明,CLL细胞保留了其起源细胞的表观遗传印记,同时显示出反映遗传亚克隆多样性的实质性患者内表观遗传异质性。这种表观遗传变异性促进了表型对微环境信号和治疗压力的适应,使得遗传上相似的亚克隆能够表现出不同的增殖和存活能力。在此背景下,表观遗传决定的累积有丝分裂(epiCMIT)评分通过量化细胞分裂相关的DNA甲基化变异性来捕获CLL细胞的增殖历史,该评分与表观遗传和遗传异质性增加、动态克隆演化以及不良临床特征相关,这与增强的演化适应性一致。
在疾病进展和治疗过程中,表观遗传状态可以动态变化,促进克隆选择、治疗耐药和复发,有时在新遗传病变出现之前或在其缺失时发生。表观遗传调节因子(如CHD2、ARID1A和SETD2)的突变进一步强化了异常的染色质状态,强调了表观遗传重编程作为CLL中可遗传且互补的克隆演化层,影响临床结局。
2.2. CLL中克隆演化的模式
CLL 例证了一种由不同克隆演化模式(稳定型、线性和分支型)塑造的恶性肿瘤,反映了白血病克隆在内在和治疗压力下适应和扩增的不同轨迹(图1)。理解这些模式对于解读治疗结局至关重要,特别是在传统CIT与靶向药物的背景下。
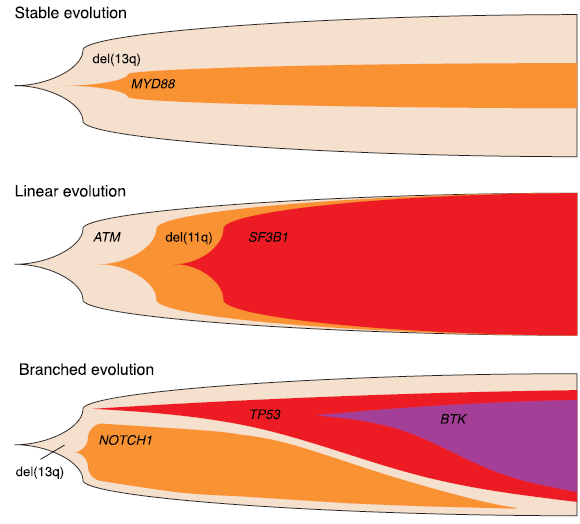
图1. 主要克隆演化模式的示意图。 稳定演化描绘了随时间推移遗传多样化最小,主导克隆持续存在而无显著选择性清除。线性演化说明了遗传改变的依次获得,其中后续克隆源自单一祖先克隆并取代其前身。分支演化显示了多个遗传上不同的亚克隆从共同祖先平行出现,反映了群体内异质性和不同的演化轨迹。
稳定型演化描述了CLL克隆组成随时间相对不变的情况。在这些案例中,创始克隆占主导地位,没有显著的新亚克隆出现(图1)。临床上,具有稳定克隆结构的患者通常表现为惰性疾病,治疗间隔期长。稳定演化更常见于具有有利预后标志物(如IGHV突变基因)的患者,这些患者甚至在特定亚组中可能实现类似于治愈的长期缓解。
当连续的亚克隆从优势克隆中出现时,就会发生线性进化,以逐步的方式相互替代。每个演化步骤通常以获得赋予生长或存活优势的额外驱动突变为标志(图1)。在FCR治疗下,由于化疗施加的强大选择压力,可以观察到线性演化模式,消除敏感克隆并允许耐药群体(通常携带TP53、ATM或RPS15突变)占主导地位。这种模式与CIT初始反应后的侵袭性疾病进展和复发相关。
分支演化是最复杂的模型,其中多个亚克隆从共同祖先分化出来并共存、竞争或合作,不同的亚克隆在不同的微环境或治疗条件下获得选择性优势(图1)。该模型在靶向治疗时代尤其相关。例如,使用BTK抑制剂如伊布替尼时,携带BTK或PLCG2突变的耐药亚克隆可以与其他竞争性克隆一起出现,导致异质性耐药机制。类似地,维奈克拉施加的选择压力有利于耐药BCL2变体的出现。与FCR不同,由于其作用机制较窄,靶向治疗通常促进分支模式,允许多种耐药通路共存。
3. 克隆演化的特殊情境
克隆演化也支撑着重要的疾病过渡阶段,如从单克隆B细胞淋巴细胞增多症(MBL)进展为CLL、Richter转化(RT)以及U-CLL与IGHV突变型CLL(M-CLL)的不同临床行为。在每种情况下,高危亚克隆的扩增通常由TME或治疗的选择压力驱动,决定了临床结局。
3.1. MBL到CLL的克隆演化
MBL是CLL的前驱状态,定义为外周血中存在少量克隆性B细胞(<5×10⁹/L),无CLL的临床症状如淋巴结肿大、器官肿大或血细胞减少。虽然大多数MBL病例保持惰性和临床静默,但一部分以每年1-2%的估计率进展为显性CLL。从MBL到CLL的演化为白血病发生和疾病进展机制提供了关键见解。
MBL中最初的克隆扩增可能源于单个B细胞中的遗传或表观遗传事件,赋予增殖或存活优势。基因组研究表明,MBL中的突变谱与CLL相似,但驱动突变的频率较低。在低计数MBL中很少检测到NOTCH1、SF3B1、TP53和ATM等基因的突变,尽管仍然罕见,但在高计数MBL和早期CLL中变得更常见。这表明遗传病变的逐步积累,早期克隆建立了MBL表型,随后的打击驱动了向症状性CLL的进展。重要的是,IGHV基因突变状态也提供信息:具有未突变IGHV基因的MBL个体比具有突变IGHV基因的MBL个体进展风险更高。
从MBL到CLL的转变不仅受内在突变影响,还受BCR信号或TME驱动的选择压力影响,包括基质相互作用、细胞因子和免疫监视。最终,适应性更强的亚克隆占主导地位,导致白血病负荷增加和CLL特征性的临床表现。临床上,区分MBL和早期CLL主要依据淋巴细胞计数和临床发现,而非仅靠生物学,强调了它们的连续性。低计数MBL通常在克隆大小上保持稳定,很少发展为显性疾病,而高计数MBL进展为需要治疗的CLL的可能性更大。通过灵敏的分子技术(如NGS)监测克隆结构,已为这种转变过程中的逐渐亚克隆选择提供了强有力的证据,如几项研究所证明。
3.1.1. U-CLL与M-CLL中的克隆演化
U-CLL和M-CLL之间的临床差异反映了不同的克隆演化模式。U-CLL的特征危更大的基因组不稳定性和更高的动态克隆演化倾向。携带未突变IGHV基因的克隆对BCR信号反应更灵敏,导致慢性激活、增殖和存活优势。随着时间的推移,U-CLL倾向于积累高危遗传改变,这些改变通常发生在选择压力(尤其是治疗)下扩增的亚克隆中。纵向研究表明,U-CLL显示出克隆主导地位的频繁转变,耐药亚克隆取代敏感克隆,这是分支演化的标志。这种动态模式解释了U-CLL中更短的TTFT、更高的RT风险以及对CIT和靶向药物更强的耐药可能性。
相比之下,具有突变IGHV基因的肿瘤表现出降低的BCR信号能力和普遍较低的增殖驱动力,因此M-CLL通常表现出更稳定的克隆结构,随时间亚克隆多样化有限。许多M-CLL患者表现出长时间不需要治疗,当进展发生时,通常由逐渐的克隆选择驱动,而非侵袭性亚克隆的快速出现。尽管TP53、SF3B1或ATM等突变可能发生在M-CLL中,但其频率显著低于U-CLL。此外,携带TP53突变的M-CLL患者在长期随访中可以在不需要治疗的情况下保持临床稳定,因此M-CLL倾向于表现为惰性演化和较低的转化风险。
治疗进一步加剧了U-CLL和M-CLL之间克隆动力学的差异。CIT(如FCR)在部分M-CLL患者中可诱导持久缓解,有时达到类似治愈的长期疾病控制。然而在U-CLL中,由于耐药克隆的快速生长,FCR反应持续时间较短。对于BTKi或BCL2i等靶向治疗,U-CLL中的克隆演化通常表现为耐药亚克隆的出现,而M-CLL患者更常维持持久反应。
3.1.2. Richter转化中的克隆演化
RT是指在潜在CLL患者中发展为侵袭性淋巴瘤,最常见的是弥漫大B细胞淋巴瘤(DLBCL)。发生在约2-10%的CLL病例中,RT是克隆演化的一个显著例子,即惰性白血病克隆获得额外的遗传和表观遗传改变,赋予更具侵袭性的表型。理解这一过程至关重要,因为RT与极差的预后和对常规治疗的耐药相关。
RT中的克隆演化可以遵循两个主要轨迹:
-
克隆相关RT:转化的淋巴瘤起源于原始的CLL克隆,因此携带相同的IGHV基因重排。
-
克隆无关RT:RT代表一个独立的新发淋巴瘤,最初的RT分歧发生在CLL诊断之前,并且RT克隆随后经历了显著的克隆扩增。
大多数病例是克隆相关的,反映了白血病克隆的真正线性进展,而10-20%是无关的,并且预后较好。在克隆相关RT中,转化是由高危遗传改变的获得驱动的,这些改变破坏了肿瘤抑制因子,激活了致癌信号,并实现了免疫逃逸。其中最常见的是TP53突变或缺失,它损害DNA损伤反应并促进基因组不稳定性。CDKN2A/B丢失导致细胞周期控制失调,而MYC激活驱动不受控制的增殖。NOTCH1和NF-κB通路的畸变进一步增强了生存信号。这些改变共同标志着从CLL的缓慢生长生物学向淋巴瘤的侵袭性行为的转变。
表观遗传重塑也在RT演化中发挥作用。染色质修饰因子如EZH2和CREBBP的改变导致转录重编程,促进淋巴瘤样表型。与此同时,TME(包括与T细胞和基质成分的相互作用)支持免疫逃逸和侵袭性克隆选择。
3.2. 评估CLL中的克隆演化:临床与研究背景
评估CLL中的克隆演化为疾病进展提供了重要见解,并可能有助于预测治疗反应。然而用于评估克隆动力学的方法在临床实践和研究背景之间存在显著差异,主要反映了技术复杂性、灵敏度和临床适用性的差异。
在临床实践中,克隆演化的评估主要侧重于识别已确立的预后和预测标志物,以指导治疗决策。应用FISH分析来识别高危患者,特别是del(17p)。常规核型分析也可揭示CK,但并非普遍推荐。在分子水平上,应用靶向NGS来检测高危突变,包括TP53、NOTCH1和SF3B1,并确定IGHV基因突变状态。虽然这些方法已标准化并整合到临床指南中,但它们仅捕获主导克隆的快照,不涉及亚克隆水平的详细表征。
相比之下,基于研究的方法采用高分辨率技术,旨在绘制CLL克隆结构的全部复杂性。全外显子测序(WES)和全基因组测序(WGS)允许无偏倚地发现编码和非编码突变。超深度NGS提供更高的灵敏度,以检测可能驱动疾病进展、复发或治疗耐药的罕见亚克隆变异。此外,单细胞测序技术通过在细胞水平上解析异质性,彻底改变了克隆演化的研究,能够重建谱系关系和演化轨迹,并揭示CLL中显著的克隆内异质性,在通过混合样本分析看似单一的克隆内共存着遗传上不同的亚群。这些技术不仅能够详细剖析个体患者体内的亚克隆组成和演化动力学,还能在疾病进展和治疗反应过程中纵向跟踪这些过程。
除了遗传改变,研究策略还扩展到表观遗传和转录组学分析,以及多组学整合,提供了关于遗传、表观遗传和转录程序如何相互作用以塑造克隆适应性的见解。纵向采样结合计算建模进一步增强了随时间推移和在不同治疗下跟踪亚克隆动态的能力。
如前所述,临床和研究评估之间的关键区别在于范围和分辨率。临床方法优先考虑可操作、标准化的标志物来指导患者管理,而研究方法旨在深入揭示CLL的演化生物学。弥合这些领域仍然至关重要,因为研究的发现(例如耐药突变的早期检测或新疗法下的克隆选择)正在逐步转化为临床相关检测,从而实现更加个性化和适应性的治疗策略。
3.3. 靶向治疗期间发生耐药的影响
以下章节将探讨导致对靶向治疗(特别是BTK和BCL2抑制剂)产生耐药的分子机制。理解这些机制至关重要,因为它们不仅解释了为什么某些治疗会随时间失效,还揭示了可用于治疗的可靶向弱点。
4. 使用共价BTK抑制剂靶向BTK
BcR信号对于CLL细胞的存活和增殖至关重要,抑制BTK是破坏BcR信号的一种高效策略。首创的共价BTK抑制剂(cBTKi)伊布替尼,即使在传统上接受CIT治疗结局较差的患者中(例如那些具有未突变IGHV基因和/或TP53畸变的患者),也显示出显著的疗效。第二代cBTKi,例如阿可替尼和泽布替尼,旨在更选择性地靶向BTK,从而减少脱靶不良事件。当与伊布替尼直接比较时,两种第二代BTKi都非常有效,同时具有更少的不良反应。最近,第三种第二代cBTKi奥布替尼已在中国获批用于治疗初治(TN)和复发/难治性(R/R)CLL。奥布替尼尚未在随机对照试验(RCT)中与其他cBTKi进行直接比较。
所有cBTKi最初都是作为连续治疗给药,要么作为单一疗法,要么与抗CD20抗体联合使用,直至进展或出现不可耐受的毒性。近年来,BTKi与BCL2抑制剂联合作为固定疗程方案的一部分。大多数接受连续cBTKi治疗的患者实现了长期缓解,原发性难治性疾病罕见,尤其是在一线治疗中。然而,在初始反应(通常持续数年)之后,相当数量的接受cBTKi治疗的患者最终会出现CLL复发。已有多种机制被用于解释cBTKi的耐药性。
4.1. 进展时的BTK/PLCG2突变
BTK和PLCG2基因中的突变是研究最广泛的cBTKi耐药机制,其中导致第481位半胱氨酸被丝氨酸取代(Cys481Ser)的BTK突变是在cBTKi治疗进展患者中最常检测到的突变。所有cBTKi都不可逆地与BTK蛋白的Cys481残基结合,因此BTK Cys481Ser突变直接影响药物结合,阻碍BTK抑制并恢复BcR信号传导。
其他BTK突变也有报道,但频率较低。具体来说,Leu528Trp、Cys481Arg和Val416Leu突变影响BTK结合和激酶功能,而Thr474Ile是一种看门人突变,破坏了对催化结构域的访问(图2)。Thr417Ile通常与Cys481Ser同时发生,主要在阿可替尼和奥布替尼治疗后检测到。另一方面,Leu528Trp在泽布替尼治疗后更常发生。
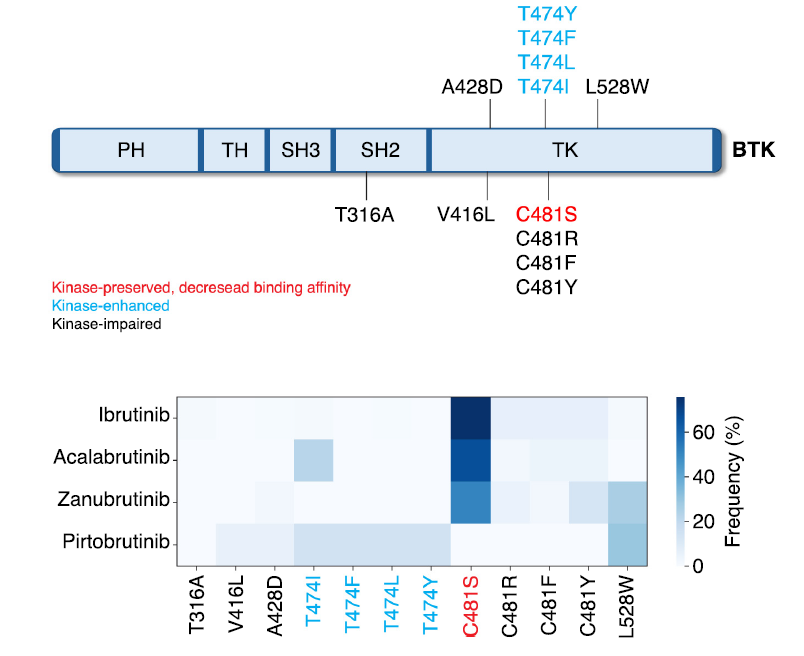
图2. BTK蛋白的线性示意图,突出了使用共价BTK抑制剂(伊布替尼、阿可替尼和泽布替尼)和非共价BTK抑制剂(匹妥布替尼)治疗后出现的最常见耐药突变。热图显示了每种抑制剂所报告的BTK突变的相对频率。
在cBTKi治疗期间CLL进展时,在少数患者中可检测到PLCG2突变。这些突变是功能获得性的,导致PLCG2蛋白的组成性激活和对上游信号传导的敏感性增加。PLCG2突变通常以低VAF检测到,并且几乎总是与BTK突变同时发生。
4.2. 持续临床缓解期间BTK/PLCG2突变的出现
多项研究报告了在接受BTKi(主要是伊布替尼)治疗但无临床进展的患者中检测到BTK/PLCG2突变。这些突变在一线治疗患者中相对罕见,但在接受伊布替尼作为R/R治疗的患者中更常发生且在更短暴露后出现。虽然携带BTK/PLCG2突变但无临床进展的患者仍可从继续BTKi治疗中获益,但预计他们比无突变的患者复发更快。在大多数研究中,从突变检测到临床复发的中位时间不到一年,尽管个体间差异显著。
4.3. BTK抑制的替代耐药机制
在BTKi治疗期间进展的相当比例患者(至少30%)中不存在BTK和PLCG2突变。此外,这些突变通常以非常低的VAF出现,质疑了其作为主要耐药机制的作用。然而需要强调的是,上述研究中的BTK/PLCG2突变是在外周血和/或骨髓中检测的,可能低估了淋巴结内的真实VAF,特别是在以淋巴结肿大加重为进展特征的患者中。尽管如此,在无BTK/PLCG2突变的病例和体外研究中已描述了BTKi耐药的潜在替代机制。这些包括:基因突变(例如EGR2、NFKBIE和CARD11基因)、CNA(例如2p和8p染色体臂的缺失)、替代通路(MAPK和Toll样受体通路)的激活以及TME相互作用(例如护士样细胞和TME激动剂介导的耐药)。
4.4. 非共价BTK抑制剂
非共价BTK抑制剂(ncBTKi)旨在克服Cys481结合位点的突变。在BRUIN研究中,目前唯一获批的ncBTKi匹妥布替尼被证明对先前接受过BTKi治疗的患者有效,无论是否存在BTK Cys481Ser突变。有趣的是,携带PLCG2突变(18例)的患者是唯一总缓解率较低的亚组。
随后,在一项纳入既往接受过cBTKi治疗患者(包括携带BTK和PLCG2突变的患者,在匹妥布替尼组中分别占37%和15%)的3期试验中,发现匹妥布替尼优于idelalisib加利妥昔单抗或苯达莫司汀加利妥昔单抗=。尽管匹妥布替尼在这个难以治疗的人群中显示出显著的疗效,但两项研究的中位PFS均不到2年。与cBTKi类似,BTK(Cys481Ser以外的BTK突变)和PLCG2突变代表了匹妥布替尼耐药的主要机制。最常检测到的BTK突变是Thr474和Leu528Trp,这些突变也可以在cBTKi治疗后检测到(图2)。
4.5. 其他正在开发的BTKi
Tirabrutinib是一种第二代cBTKi,也已被测试,但尚未在CLL中获批。在一项对9名替拉鲁替尼治疗后进展患者的研究中,BTK Cys481Ser和Leu528Trp是最常检测到的变异(各4/9例,其中2例同时发生),其次是Thr474Ile(3例,均与Cys481Ser同时发生)。
ncBTKi nembrutinib在针对携带BTK Cys481Ser突变患者的1/2期BELLWAVE-001和BELLWAVE-003研究中显示出有希望的疗效,目前正在3期试验中于TN和R/R背景下进行测试。有趣的是,在BELLWAVE-003研究中接受nembrutinib治疗的92名患者中,没有人出现新的BTK突变,只有2人获得了PLCG2的耐药突变。另外两种ncBTKi,vecabrutinib和fenebrutinib,也已在1期试验中使用,但它们在CLL中的开发已停止。目前缺乏关于这些ncBTKi耐药机制的报道。
4.6. 靶向BTK的新方法
靶向BTK的最新分子包括BTK降解剂、双重共价/可逆BTKi以及双重LYN/BTK抑制剂birelintinib(即DZD8586)。BTK降解剂使用新型蛋白水解靶向嵌合体技术,通过泛素-蛋白酶体系统选择性降解BTK。几种分子(BGB-16673、NX-2127和NX-5948)目前正在CLL中进行研究,即使在ncBTKi进展后的患者中也显示出有希望的结果。另一方面,双重共价/可逆BTKi LP-168和DZD8586使用不同的机制来克服BTK突变。前者共价结合野生型BTK,非共价结合BTK耐药变异体,而后者非共价抑制BTK和LYN。这些药物正在进行临床研究,这将确定它们在CLL治疗武器库中的作用。
尽管已有报道称在接受BTK降解剂治疗的患者中出现新的BTK突变(例如BTK Ala428Asp),但尚缺乏关于BTK降解剂和双重共价/可逆BTKi耐药机制的综合研究。
5. 基于BCL2抑制的靶向治疗
BCL2家族蛋白,特别是BCL2,在CLL细胞存活中发挥关键作用。CLL细胞在微环境刺激下上调BCL2,这一机制对于将凋亡平衡转向存活至关重要。作为一个关键靶点,在过去十年中,已经开发了不同的化合物来促进细胞凋亡。最初在临床试验中测试的那些选择性较差,通过在临床浓度下同时抑制BCL2和BCLxL,它们与更具挑战性的安全性特征相关,特别是由于BCLxL抑制相关的血小板减少症。专注于选择性BCL2抑制,随后开发了维奈克拉(ABT199),并在初治和复发/难治性CLL的1-3期临床试验中进行了测试。维奈克拉已获批并目前在临床实践中可用于CLL治疗,可单独使用或与抗CD20单克隆抗体(即奥妥珠单抗或利妥昔单抗)和/或cBTKi(伊布替尼或阿可替尼)联合使用。
维奈克拉作为BH3模拟物,与BCL2的疏水沟结合;结合后,促凋亡的BH3蛋白被置换,使平衡转向凋亡。维奈克拉单独和/或与抗CD20单克隆抗体或BTKi联合使用被证明对绝大多数CLL患者有效,并能实现深度且持久环节。尽管如此,随着观察时间的延长,耐药的出现也成为疗效缺乏和治疗中断的潜在原因。
5.1. BCL2抑制的耐药机制
维奈克拉耐药的潜在机制相当异质性,并且在单个患者的进展时间点识别出不止一种机制通常是常态。值得注意的是,关于维奈克拉耐药的数据来源于接受连续治疗的患者(作为维奈克拉最初的给药方式,与BTKi一致),而关于固定疗程联合治疗后出现耐药的数据仍然很少。对维奈克拉的原发性耐药,尤其是在初治背景下,是极其罕见的事件,主要发生在早期1-2期试验中入组的大量预处理患者群体中。在当前的治疗场景中,继发性耐药(即初始反应后失去反应)是最常见的事件。维奈克拉耐药机制可能是细胞内在的和细胞外在的(由微环境相互作用促进),但在大多数情况下这种区分是人为的,因为两者都有助于全面临床耐药性的发展。
在早期临床试验中,在接受维奈克拉治疗后复发的患者中发现了BCL2点突变的发生。可能发生几种突变,其中Gly101Val是最常见的。BCL2突变的模式支持瘤内异质性,因为在复发时通常存在多个携带不同突变的克隆。Gly101Val突变与维奈克拉与BH3沟的结合受损相关,导致BCL2抑制不理想。突变的BCL2仍保留其抗凋亡功能,有效结合促凋亡家族成员,如BIM、BAX和BAK。BCL2突变通常在接受连续维奈克拉治疗的患者中被记录,并且往往在治疗至少12-24个月后发生。
尽管最初在维奈克拉耐药病例中发现了BCL2突变,但最常见的维奈克拉耐药机制与其他抗凋亡BCL2家族蛋白的上调相关,迄今为止最强的证据是MCL1的上调。MCL1上调可以通过几种机制实现,既有细胞内在的,也有细胞外在的。微环境刺激如IL4、CD40L和BAFF可能直接或通过NF-κB激活诱导MCL1上调。MCL1活性可通过超磷酸化增加,而维奈克拉治疗后的染色体不稳定性可能触发1号染色体事件的扩增。
耐药病例中促凋亡基因的变化很少被检测到,例如在个别病例中记录了BAX或NOXA的亚克隆丢失。在少数病例中报道了导致截短、无功能蛋白的BAX剪接改变(图3)。
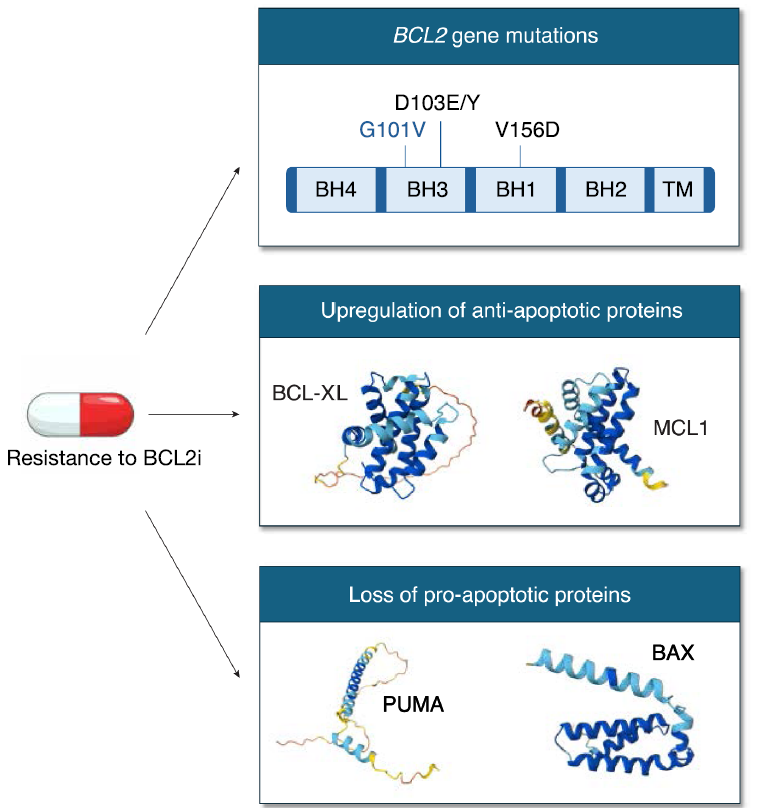
图3. BCL2抑制剂靶向治疗的耐药机制。 蛋白质结构来自AlphaFold蛋白质结构数据库。颜色表示模型置信度,从非常高(蓝色)到非常低(橙色)。
5.2. 克隆演化和表观遗传学在维奈克拉耐药中的作用
克隆发展和遗传不稳定性可能是导致CLL中维奈克拉耐药的潜在机制,但尚未识别出独特的克隆演化模式,病例表现为线性、分支、发散和趋同演化。在维奈克拉治疗期间,CNA和点突变的数量增加,表明在持续BCL2抑制的压力下,适应性最强的克隆发生扩增。NOTCH1 突变和 CDKN2A/B 缺失有利于 CLL 细胞存活和增殖,可以抵消凋亡启动。特别是,CDKN2A/B缺失与接受维奈克拉治疗的患者中的RT相关。BTG1突变在维奈克拉治疗后的复发中反复富集,与治疗驱动的克隆选择一致。BTG1功能丧失可能通过损害抗增殖和肿瘤抑制程序,降低细胞对BCL2介导的凋亡的依赖性,从而促成耐药。这些改变通常与其他遗传病变(例如CDKN2A/B丢失)同时发生,共同在BCL2抑制下促进存活和增殖。
除了遗传改变,表观遗传修饰也与维奈克拉耐药相关。从血液恶性肿瘤和维奈克拉耐药患者收集的耐药细胞系和原始样本中,识别出PUMA(一种关键的促凋亡蛋白)的表观遗传沉默是与临床获得性维奈克拉耐药时基线相比的显著变化]。PUMA下调和丢失导致细胞代谢增加,这得到了基础和最大呼吸水平以及氧化磷酸化增加的支持。
除了细胞内在的耐药机制外,初步证据表明,微环境保护也可以通过CD40-CD40L、BAFF/APRIL信号通路以及细胞因子和趋化因子的释放(包括IL4、IL6、IFN、CXCL12和CXCL13)以及整合素和粘附分子来促进耐药。
5.3. 新型BCL2抑制剂
在过去几年中,开发了新型BCL2抑制剂,目前正在临床试验中使用,其中进展最快的是索托克拉和利沙托克拉。索托克拉是一种更具选择性和更强效的BCL2抑制剂,目前正在3期临床试验中开发。临床前数据表明,在几种B细胞恶性肿瘤模型中,与维奈克拉相比,索托克拉显示出更强的细胞毒活性和更深入的肿瘤生长抑制作用。更重要的是,在体外,索托克拉能够有效抑制对维奈克拉耐药的BCL2变异体,包括Gly101Val突变。BCL2wt和BCL2G101V与索托克拉结合的晶体结构证实,索托克拉结合野生型和突变型BCL2的P2口袋,这种新的结合模式可能解释了其在BCL2 Gly101Val形式中也具有更强活性的原因。
利沙托克拉是一种新型BCL2抑制剂,对BCL2蛋白具有高结合亲和力和选择性,目前正在3期试验中研究。尚无关于利沙托克拉在对维奈克拉产生耐药的患者中疗效的数据。导致索托克拉或利沙托克拉耐药的机制仍远未阐明。
6. 未来方向与结论性评述
在过去十年中,基因组分析和靶向治疗的重大进展从根本上改变了对CLL的理解和管理。然而正如本综述所强调的,克隆演化仍然是驱动疾病进展和治疗耐药的核心生物学过程。尽管使用BTKi和BCL2i获得了令人印象深刻的缓解率,但CLL对大多数患者来说仍然无法治愈,这强调了对耐药和复发机制进行更深入机制研究的必要性。
尽管国际指南通常不推荐检测耐药变异体(如BTK、PLCG2和BCL2中的突变),但诊断检测正成为常规临床实践中越来越常见的部分。正如欧洲CLL研究倡议(ERIC)最近建议的,当预期检测结果会影响患者管理时(例如,在开始新线治疗之前、在既往暴露于cBTKi后使用ncBTKi治疗时,或考虑使用BCL2i或BTKi再治疗时),应进行诊断检测。
此外,将耐药机制的识别视为涉及靶向治疗的临床试验的关键组成部分。未来的研究必须采用综合性的高分辨率方法,以完全捕获CLL演化的复杂性。结合基因组学、转录组学和蛋白质组学的多组学策略对于剖析遗传病变如何与转录程序和信号网络相互作用以塑造克隆适应性至关重要。特别是,蛋白质组基因组分析可以通过直接询问从单纯DNA测序无法预测的通路激活、翻译后修饰和适应性信号重连,来弥合基因型与表型之间的差距。
单细胞测序技术也可以在定义耐药的细胞结构方面发挥重要作用。通过在单个细胞水平上解析异质性,这些方法能够重建演化轨迹、识别罕见的耐药微克隆,并描绘亚克隆之间的协同相互作用。单细胞基因组学与单细胞转录组学的整合将进一步阐明遗传相似的克隆如何在治疗压力下发生功能分化。空间组学方法代表了另一个前沿领域,允许研究解剖学微环境(尤其是淋巴结)如何塑造克隆选择、药物耐受性和微环境介导的耐药,而这些通常被外周血分析低估。
重要的是,未来的研究必须解决数据仍然稀少的新兴临床情况。在接受固定疗程伊布替尼-维奈克拉治疗后复发的患者中,BTK、PLCG2和BCL2突变似乎基本上不存在,这意味着治疗持续时间和药物顺序可能影响克隆选择,尽管现有数据仍然有限。同样,对BTK和BCL2抑制均耐药的双重难治性CLL的生物学是一个关键的未满足需求,因为这些患者的结局特别差。对此类病例的系统性分子表征对于指导合理的序贯和联合策略至关重要。此外,新型免疫治疗方法,包括双特异性抗体和CAR-T细胞治疗,引入了新的选择压力。
总之,CLL 例证了由动态和适应性克隆进化驱动的恶性肿瘤。利用新一代多组学、单细胞和空间技术,纵向和跨治疗方式应用,将是解开耐药性生物学的关键。将这些见解转化为临床可操作的诊断工具,为真正个性化、进化知情的治疗提供了希望,最终改善 CLL 患者的长期结局。
参考文献
Semin Cancer Biol . 2026 Apr 17:122:16-29. doi: 10.1016/j.semcancer.2026.04.001.

