Immunity:疾病相关小胶质细胞和疾病炎症相关巨噬细胞在衰老和神经退行性变中的双重发育特征
时间:2022-10-23 12:00:38 热度:37.1℃ 作者:网络
导读:
大脑内巨噬细胞群包括实质小胶质细胞、边界相关巨噬细胞和募集的单核细胞衍生细胞。这些细胞共同控制大脑的发育和稳态,也与衰老的发病机制和神经退行性变有关。然而每种细胞群体在不同背景下的表型、定位和功能尚未解决。近期《Immunity》期刊发表了题为《Dual ontogeny of disease-associated microglia and disease inflammatory macrophages in aging and neurodegeneration》的研究论文,作者团队建立了一个小鼠脑髓系scRNA-seq整合来系统地描述脑巨噬细胞群。结果发现,先前在小鼠阿尔茨海默病模型中发现的疾病相关小胶质细胞(DAM)群体实际上包括两种在个体发育和功能上不同的细胞谱系:即表达神经保护信号的胚胎来源的触发受体骨髓细胞2(TREM2)依赖的DAM,以及在衰老过程中积累在大脑中的单核细胞来源的TREM2表达的疾病炎性巨噬细胞(DIMs)。而且这两个不同的细胞种群在人脑中似乎也是保守的。在这里,作者建立了脑髓系细胞在发育、内稳态和疾病中的异质性本体发育解析模型,并确定了治疗神经退行性变的细胞靶点。

1.M-Verse:生成通用集成以识别跨多个数据集的鼠脑巨噬细胞亚群
鉴于不同的单细胞RNA测序(scRNA-seq)研究对小鼠脑中巨噬细胞亚群的研究方法和结论各不相同,作者团队首先旨在整合其他研究中收集的六个数据集和使用Seurat产生的两个内部数据集(Silvin IS,Silvin C1)。为了确保所产生的整合—作者团队将其命名为“M-Verse”(图1A),并选择已发表的数据集来涵盖中枢神经系统内的一系列广泛情况:一种侧重于比较幼年和成年小鼠的小胶质细胞和神经元、中间神经元、少突胶质细胞和星形胶质细胞(Zeisel);另一种追踪了胚胎到衰老小胶质细胞的转变,其时间点分别在E14.5、P4、P5、P30、P100和P540(Hammond);还有一种是神经退行性变中的小胶质细胞,包含了对DAM的首次描述(Keren-Shaul)。来自Van Hove的数据集还为分析增加了空间维度,因为它包括来自不同脑区的细胞:脉络丛、硬脑膜、硬脑膜下区域(SDMs)和脑实质(全脑)。作者团队的数据集包括从胚胎(E12.5、E16.5和E18.5)、出生后早期(P7、P14)和成年(P60)小鼠大脑中分离的小胶质细胞,以及成年小鼠(P60)的CD45+巨噬细胞。作者团队期望将这些不同的数据集放在一起从而构建产生一个强大的工具来解析小鼠脑巨噬细胞的异质性。
MVerse(https://macroverse.gustaveroussy.fr/2021_M-VERSE)允许作者团队提取在所有数据集中检测到的14,794个共同基因的表达数据。通过比较这些数据集,作者团队生成了发育中和成年小鼠大脑中髓系细胞的无偏全局图并确定了图中的五个区域(图1A):一个对应于非髓系细胞(包括一个对应于神经元的集群和一个对应于中间神经元、少突胶质细胞和星形胶质细胞的集群);一个对应于T细胞和NK细胞;一个富含表达发育小胶质细胞特征的细胞;一个富含具有成熟小胶质细胞特征的细胞;以及第五个含有非小胶质细胞的髓系细胞。
当作者团队将每项研究的数据分别投射到M-Verse(图1B)时,他们发现Van Hove数据集富含髓系细胞,这可能是由于脑膜和脉络丛区域的scRNA-seq影响。Silvin C1和Hammond数据集是仅有的两个包含胚胎和出生后早期小胶质细胞的数据集,并且都产生了一个明确的发育期小胶质细胞特征簇。Zeisel数据集包含高比例的非髓系细胞(分化神经元[Tubba2,sox11],中间神经元[Slc32a1,Pnoc],少突胶质细胞[Hapln2,Opol1]和星形胶质细胞[Slc1a2,Slc1a3],在M-Verse的神经元区域附近清晰可见 [Stmn1,Tub2b] (图1B))。尽管如此,Zeisel数据集中的小胶质细胞的表达谱与其他数据集的小胶质细胞完全重叠,表明整合是成功的。值得注意的是,Zeisel数据集还包含一些聚集在发育特征区域的小胶质细胞,这可能是由于它们使用了青少年期或幼年小鼠(从21天到1个月)。同样,作者团队在Hammond、Silvin C1和Silvin IS数据集(图1A、1B)中观察到了60-100日龄小鼠发育小胶质细胞特征区的少量细胞,并且与老年WT小鼠(Keren-Shaul 和 Van Hove)(图 1B)相比,患有神经退行性疾病的老年小鼠该区域的细胞增加。这表明在这些神经退行性疾病模型中,一些小胶质细胞可能获得类似发育的程序。
2.使用小鼠脑巨噬细胞的定向索引排序数据完善M-Verse
scRNA-seq方法提供了宝贵的数据,通过对少量细胞的深入分析来补充广泛的分析的结果,可以促进对细胞亚群的更精确识别。因此,作者团队接下来使用Seurat V2(图1A中红色显示)分析了Silvin IS数据集(该数据集结合了蛋白质表达信息和深度基因表达测量信息)。该数据集包含来自213个成年鼠脑 CD45+Ly6C- F4/80+CD11b+巨噬细胞的信息并使用Smart-Seq2方法生成。该方法能够测量与10X Genomics类似数量的基因,而且每个细胞具有更多的片段读取数量。这样就能够与IS相结合以分离具有已知特征(例如,定义的大小、颗粒和选定的表达标记)的单个细胞。为了明确识别传统门控小胶质细胞群中的边界相关巨噬细胞(BAMs),作者团队使用来自2月龄的 Lyz2GFP小鼠的细胞,其中含有GFP标记表达溶菌酶的BAMs(而不是小胶质细胞)。然后使用Seurat V2和Phenograph方法对213个测序细胞进行了聚类,并根据它们的scRNA-seq表达谱确定三个集群:一个大集群1和两个小集群2和3。
为了进一步表征这3个集群,作者团队通过流式细胞技术获得了各组“指标”表面分子的表达。结果发现次要集群2和3表达更高的CD45和Lyz2GFP,而主要集群1表达中等量的CD45、F4/80、CD11b和Ly6C及少量Lyz2GFP(一种类似小胶质细胞的表型)。为了更好地理解集群2 和3的组成,作者使用之前的参数独立于其它细胞重新分析了Lyz2GFP+细胞(BAMs)。基于基因表达谱,集群2 被解析为两个子集群并将其注释为2a和2b(图 1C,左图)。然后根据流式细胞仪数据可以发现集群2a更像小胶质细胞(集群 1),而集群2b与集群1 明显分离,特别是其中表达大量的MHCII 蛋白。为了进一步研究集群的特性,作者团队使用了差异表达基因 (DEG) 分析,结果表明集群1对应于小胶质细胞(Tmem119 的高表达)。尽管集群2的特征是BAM标记物Mrc1 (Cd206) 的高表达为特征,但集群2b在RNA和蛋白质量上表现出比集群2a更高的H2-Ab1、H2-Aa 和 Cd74 表达(图 1D)。最后,集群3代表一个独特的细胞群,其特征是表达Cd11a、Cd44、Cd300e、Plac8、Nr4a1、Ear2 和 Fcgr4(图 1D)。其中Cd11a 的RNA 和 CD11a 蛋白在集群3细胞之间存在相关性(图 1D)。
然后作者团队使用来自IS scRNA-seq (Silvin IS) 的新的深度数据为M-Verse中的每个细胞群生成可靠的特征。这使作者能够确认M-Verse中的CD11a+(集群 3)和 CD206+巨噬细胞(集群 2a 和 2b)与小胶质细胞(集群 1)明显聚集(图 1C)。然后再将这些特征投射到M-Verse上,以扩展和验证它们在其他数据集上的异质性。CD11a+、CD206+MHCII-和CD206+MHCII+巨噬细胞特征的探测使作者团队能够突出特定髓系细胞区域内包含表达相似基因模式的细胞数量(图 1E)。通过关注这些区域(图 1F),作者团队确定了所有数据集共有的BAMs对应的区域(图 1G)。这种方法也能够更好地分辨其他已发表研究中的细胞身份:当作者使用Van Hove脉络丛BAM (CPepiBAM) 标记来研究其包含脉络丛细胞的数据集区域时,发现表达此特征的细胞实际上是主要位于由M-Verse定义的成熟和发育的小胶质细胞区域内(图 1H)。因此,在最初的研究中被认为是BAMs的细胞很可能是小胶质细胞:已发表的脉络丛‘BAM’表达转录因子Sall1的结果也进一步支持了这一观点。因为此前人们发现该转录因子在小鼠中枢神经系统内的脑巨噬细胞中只有在小胶质细胞中表达。
总之,整合使用不同技术生成的六个脑巨噬细胞scRNA-seq数据集使作者团队能够对发育中和成年小鼠大脑进行全局交叉比较。并通过进一步应用IS单细胞分析提高M-Verse的分辨率后确定了一套共识的mRNA标记转录物,从而能够可靠和准确地识别跨多个生命阶段的小鼠小胶质细胞和脑巨噬细胞群。
3.M-Verse定向分析解决了BAMs的起源问题
基于M-Verse和标志性差异表达基因(DEG)编码蛋白的分析,作者团队设计了一种严格的流式细胞术门控策略来识别上述定义的脑巨噬细胞群。这些标记基因和蛋白质包括 Mrc1(编码 CD206)、Itgal(编码 CD11a)和 H2-Ab1(编码 MHCII)(图 1D)。作者首先评估了整个年轻成年小鼠脑(大脑、软脑膜、蛛网膜和硬脑膜)、去除脑膜的大脑(大脑和软脑膜)和单独的脑膜(蛛网膜和硬脑膜)的细胞悬液。正如预期的那样, CD11a+和 CD206+MHCII+巨噬细胞在脑膜部分中富集,而只有脑的部分则CD206+MHCII-细胞富集(图1I)。因此,这种基于综合分析确定标志DEGs的门控策略,使作者团队能够准确识别小胶质细胞和差异表达MHCII的 CD206+BAMs的两个子集,以及表达整合素CD11a的巨噬细胞子集。
建立这些基线后,作者使用最近描述的Ms4a3命运映射模型跟踪血液单核细胞对细胞群的贡献:不到 1% 的小胶质细胞被标记,而超过87% 的CD11a+巨噬细胞被标记,这表明后者主要是单核细胞衍生的。CD206+MHCII+巨噬细胞的单核细胞贡献率(20%)高于CD206+MHCII-巨噬细胞(<5%)(图1J)。然而,作者团队也观察到CD206+MHCII+单核细胞来源的细胞(26%)和小胶质样单核细胞来源的细胞(2%-3%)的百分比在大于6月龄的小鼠中略高(图1K)。总之这些结果表明,与血管周围巨噬细胞相比,脑膜CD206+MHCII+巨噬细胞具有更高的单核细胞成分,因此被替代的程度比先前认为的更大。

图1 M-Verse 鼠脑巨噬细胞异质性的整体共识
4.DAM群体内的表型异质性
DAM是一组CNS巨噬细胞,被认为在阿尔茨海默病 (AD) 小鼠模型中发挥保护作用,也存在于人类 AD 患者的大脑中。另一项研究表明,DAM也会在老年小鼠的大脑中积累,但它们是否具有保护作用仍存在争议。最初使用scRNA-seq分析确定DAM高表达Cd11c、Csf1和Cd9,低表达Tmem119、P2ry12和Cx3cr1,并取决于髓样细胞2(TREM2)上表达的触发受体。目前,DAM的本体发育仍难以捉摸,而且最近的研究得出了相互矛盾的结论--即它们可能与BAMs或单核细胞有关。
因此,作者团队试图通过M-Verse研究DAM在整个发育、成年、衰老和神经退行性变的扩展大脑巨噬细胞群体中的性质。作者将Keren-Shaul 的DAM基因表达特征投影到具有M-Verse共同坐标的Keren-Shaul 数据集上,发现高表达DAM基因特征的细胞位于两个不同的区域:一些位于发育小胶质细胞区域(DAM) ,另一些位于成熟的小胶质细胞区 (图 2A)。由于这些细胞的炎症基因高表达,因此作者将其定义为疾病炎症巨噬细胞 (DIMs)。为了评估表达DAM特征的这两个亚群是否同样依赖于TREM2,作者团队比较了它们在老年C57BL/6 WT、5XFAD(AD 模型)、TREM2-/- 和 5XFAD-TREM2-/-小鼠大脑中来自Keren-Shaul数据集的相对丰度。结果发现,AD-TREM2-/-小鼠大脑皮层中DAM细胞的比例约为AD小鼠的一半,而所有TREM2-/-小鼠的DIMs比例显著增加(图 2B)。在RNA水平上,两个群体都高度表达Trem2(图 2C),但在其他标记的表达上存在差异:与 DIMs相比,DAM的P2ry12 和Cx3cr1表达相对较低,而 Itgax (Cd11c)的表达较高(图 2C)。因此,M-Verse 结合两个区域内细胞分子比较,不仅能够识别两个细胞群共享 DAM 标志基因表达状况,而且也能独特地将基因表达状况与其一致的分子功能差异描述出来。这些数据强烈表明DAM而非DIMs代表了Keren-Shaul描述的真正DAM。
5.真正的成熟DAM显示出与YAM重叠的基因表达特征
然后,作者接下来又研究了是否可以在用于生成 M-Verse的其他数据集中检测到DAM细胞,并发现其也存在于包含来自APP-PS1 AD小鼠模型细胞的Van Hove数据集中(图 2D)。在这两个数据集中,表达DAM特征的细胞都包含在发育中的小胶质细胞区域内,暗示成熟真正的DAM与典型的胚胎小胶质细胞共享相同的基因表达模式。因此,作者进一步进行了DEG分析,比较了两个数据集中的DAM 和 DIMs,试图定义一组在DAM中表达量更高并能代表其真正特定特征的基因(图 2E)。结果在两个数据集中发现了DAM特异性高表达的8个基因:Dkk2、Fabp5、Gpnmb、Igf1、Itgax、Mamdc2、Spp1和Gm1673。然后作者团队将这些特征基因的平均表达量投射到M-Verse内的两个数据集(Keren-Shaul 和 Van Hove 叠加)上以可视化其表达特异性(图 2F),并评估它们在这些研究中使用的不同小鼠中的比例(图 2G)。来自 Silvin C1 和 Ham-mond 数据集(WT 小鼠)的细胞出现在DAM区域中(图 2H),但在这种情况下,也对应于来自胚胎(E12.5、E14.5 和 E18.5)和出生后早期(P4、P5 和 P7)时间点的细胞(图 2I)。
总之作者的分析表明,成年DAM与非AD小鼠的发育(胎儿和出生后早期)小胶质细胞具有共同的基因表达特征,包括整合素Itgax (CD11c) 的表达,已经被证明在名为P7 CD11c+的小胶质细胞子集上表达。为了了解这两种单独描述的细胞类型的可能关系,作者团队将P7 CD11c+转录组特征投射到M-Verse 上,并观察到与Keren-Shaul数据集的DAM特征显著重叠(图 2J 和 2A)。并且DAM保守特征中七个基因中的五个(Dkk2、Gpnmb、Igf1、Itgax 和 Spp1)也在Hagemeyer 和 Wlodarczyk 数据集的P7 CD11c+细胞中表达(图 2K),以及在胚胎时间点的Hammond 和 Sil-vin C1 数据集中表达(图 2H 和 2J)。因此,在神经退行性疾病中看到的DAM保守特征与健康野生型小鼠胚胎和新生儿大脑中存在的一组细胞非常相似。基于这些观察结果,作者团队建议将CD11c+P7小胶质细胞称为青年相关小胶质细胞 (YAM)。
接下来,作者团队想更好地了解这两个具有相似基因表达特征但出现在截然不同环境中的小胶质细胞亚群之间的异同。因此他们对来自Silvin C1和Hammond数据集的YAM以及来自Keren-Shaul和Van Hove数据集的DAM之间进行了DEGs比较路径分析。YAM和DAM均表达大量与吞噬作用、自噬和线粒体代谢相关的基因,但是YAM额外表达了与脂肪酸、脯氨酸和谷氨酸有关的基因,而DAM 则单独表达与抗炎反应相关的基因(图 2L)。因此,成熟DAM似乎经历了类似胎儿的重编程,转录融合为胚胎和新生儿YAM。然而,尽管YAM和DAM具有共同的基因表达模式,但前者在胚胎发生和出生后早期自然出现,代表与大脑发育相关的小胶质细胞群;而DAM则相反,在神经退行性变的背景下出现,可能参与恢复大脑稳态。
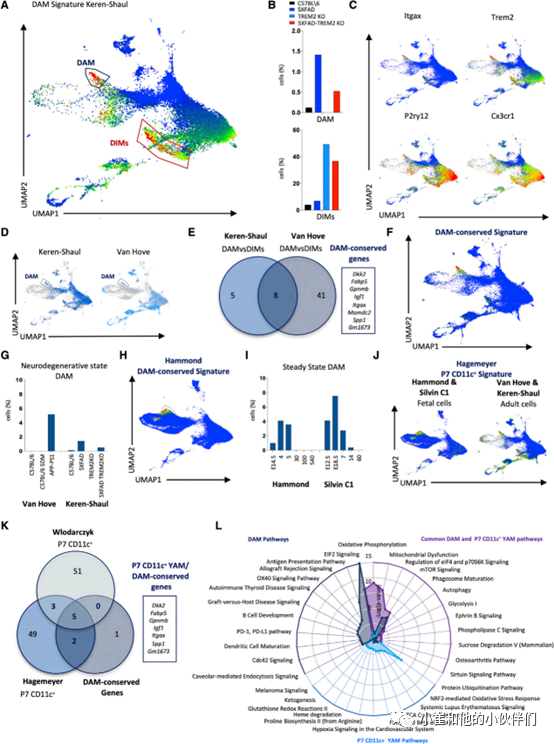
图 2 DAM 特征包括两个不同的细胞群
6.DIMs在衰老过程中出现并在神经退行性疾病中发展
然后,作者团队继续探索了DIM群体的性质并首先在小鼠神经退行性变过程中检测到了DAM的存在(图 2A)。当检查其他scRNA-seq数据集时,他们也在 Hammond、Keren-Shaul和Van Hove数据集中检测到健康老年小鼠的 DIMs(图 3A)。为了了解这些细胞在小鼠生命周期中出现的动力学,作者团队在Hammond 和Silvin C1数据集中量化了它们在不同时间点的相对丰度:DIMs 在出生后出现,并在一岁以上的小鼠中急剧增加(图 3B)。在Van Hove和Keren-Shaul数据集研究的神经退行性状态中,与年龄匹配的 WT 小鼠相比,DIMs在疾病小鼠模型中更为丰富(图 3C)。令人惊奇的是与DAM相比,Keren-Shaul数据集中的AD-TREM2-/- 小鼠的DIMs比例急剧增加,显示了它们的TREM2独立性(图 2B、3C)。通过比较来自Hammond、Keren-Shaul和Van Hove 数据集的基因表达数据,作者团队随后建立了一个DIM保守标记特征,包含了与促炎反应和免疫激活相关的基因,包括Il1a、Il1b、Tnf、Nfkbia、Cd49f、Cd54和 Cd83(图 3D、3E)。与M-Verse的其它区域相比,此特征在DIM 区域中富集。因此,鉴于炎症相关基因的高表达及其在神经退行性疾病背景下比例的增加,作者团队将这一细胞群体命名为表达TREM2(图2C)但不依赖于TREM2(图2B、3C)的炎性巨噬细胞(DIMs)。通过对DIMs与DAM专属表达的基因进行通路分析,作者观察到DIMs表达许多涉及炎症相关通路的基因,其中包括Tnf, Il6, 和Il1,以及Toll样受体信号和产生一氧化氮(NO)及活性氧(ROS)(图3F),与潜在的促炎状态一致。然而,作者团队还观察到一些免疫抑制途径,包括IL-10信号传导、糖皮质激素受体信号传导和过氧化物酶体增殖物激活受体(PPAR)信号传导途径(图 3F),这主要是由于 Ccr5、Il10ra、Tnf、Hsp90aa1、Cited2、 Icam1、Ccl2、Stat1、Fcgr1a 和 Adbr2等基因的表达可能反映了一个整合炎症环境的反馈回路。然而,作者并没有观察到免疫抑制细胞因子和趋化因子编码基因的显著表达。
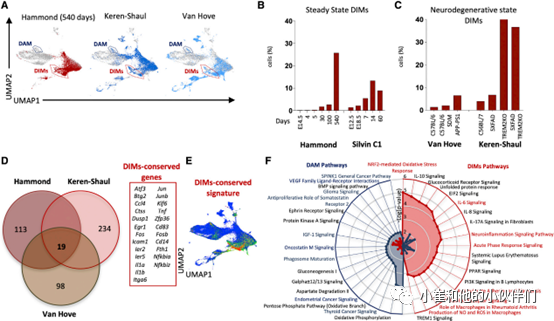
图 3 DIMs在衰老和AD小鼠模型中的比例增加
7.脑部炎症诱导单核细胞来源的CD83+DIMs的积累
除了以小鼠神经变性为特征的炎症环境外,作者团队还想了解当CNS在其他情况下发生炎症时,是否会出现DAM/DIMs样细胞。在人类中,颅骨电离辐射暴露会导致进行性认知功能障碍和神经炎症,这与体内促炎细胞因子如TNF-a、IL-1a 和IL-6的释放有关。小鼠中耗竭小胶质细胞后辐射照射和同源骨髓(BM)转移导致单核细胞渗入大脑并产生类似小胶质细胞的细胞。为了联系这些观察结果,作者团队研究了电离辐射后大脑中单核细胞衍生的巨噬细胞和小胶质细胞样细胞的存在,以了解它们是否可能与DIMs或DAM有关。通过照射1月龄的C57BL/6 WT小鼠并用Ms4a3CreRosaTomato小鼠的细胞重建它们的BM来产生BM嵌合体,然后在5个月或10个月后分析它们的大脑。移植后,Ms4a3-TdT+单核细胞的数量超过96%,但在稳定状态下,即使在重建后10个月,只有不到25%的脑巨噬细胞(根据CD45、CD11b、F4/80的表达和低表达Ly6C)表达Ms4a3CreRosaTomato。相反,照射后单核细胞来源的巨噬细胞显著增加,在随后的时间点达到20%以上。与非单核细胞衍生的巨噬细胞相比,这些Ms4a3-TdT+巨噬细胞表达更多的F4/80和CD83 以及更少的CD206。然后通过显微镜观察作者确认了大脑中存在Ms4a3-TdT+巨噬细胞。这表明Ms4a3-TdT+细胞也表达IBA1+并且分布于不同的脑区。
令作者团队特别兴奋的是照射后脑内Ms4a3-TdT+巨噬细胞上CD83的高表达,因为CD83也是M-Verse在DIMs中特异性高表达的基因之一。因此,作者团队通过透射电子显微镜 (TEM) 和 H&E 染色对CD83+Ms4a3-TdT+巨噬细胞、CD83+Ms4a3-TdT-小胶质细胞和CD83-Ms4a3-TdT-小胶质细胞进行分类和比较。结果发现与小胶质细胞相比,CD83+Ms4a3-TdT+巨噬细胞更大并且包含更多的细胞内囊泡,这可能对应于TEM可见的脂滴。作者还对这些种群进行了批量RNA测序以比较它们的基因表达谱并了解它们与DAM和DIMs的关系(图 4A)。根据CD45、F4/80、CD11b、Ly6C、CD83、和 Ms4a3-TdT(图 4B 所示的门控策略)分类:大约30%的脑巨噬细胞对应于CD83-Ms4a3-TdT-小胶质细胞,30%对应CD83+Ms4a3-TdT-小胶质细胞,40%对应于CD83+Ms4a3-TdT+巨噬细胞(图 4C)。作者团队首先使用基于M-Verse建立的更广泛的DIM基因特征的基因集富集分析(GSEA)研究这三个群体中的任何一个是否对应于RNA水平的DIMs。GSEA 在CD83+Ms4a3-TdT+巨噬细胞群中显示出明显的 DIM 特征富集(图 4D)。将此扩展到蛋白质含量分析,与 CD83-Ms4a3-TdT-小胶质细胞相比,CD83+Ms4a3-TdT+巨噬细胞表达的 CD11c 和 CD11b 量相似,但其表面的P2RY12 量略低,MHCII 量更高(图4E)。68个基因在CD83+Ms4a3-TdT+巨噬细胞和CD83+Ms4a3-TdT-小胶质细胞之间的表达量存在显著差异,包括 Axl、Irf7 和 Clec12a 以及参与神经炎症信号通路的基因,它们在CD83+Ms4a3-TdT+DIMs中被独特地上调。这些结果表明,照射后脑单核细胞衍生的巨噬细胞的特征在于其表面的CD83和F4/80表达较高,并表达与M-Verse中定义的DIM群体相对应的基因。
最后,作者团队使用 Ms4a3CreRosaTomato模型来监测2月龄和大于6月龄的小鼠大脑中DIMs的数量,并确认它们在衰老过程中的积累。使用类似的门控策略(图 4F)结果发现与成年小鼠(>6 月龄)相比,年轻成年鼠(2月龄)巨噬细胞(图 4F 和 4G)中 DIM 比例显著增加。相比之下,CD83-Ms4a3-TdT-和CD83+Ms4a3-TdT-小胶质细胞的比例在此期间没有改变。这些DIMs定位于海马、丘脑和皮层中(图 4H)。总之,DIMs 是单核细胞衍生的巨噬细胞,随着年龄的增长在大脑中积聚。
8.单核细胞来源的DIMs和YS来源的DAM在5XFAD小鼠脑内累积
然后作者团队使用流式细胞技术评估了12月龄的C57BL/6-Ms4a3CreRosaTomato、5XFAD- Ms4a3CreRosaTomato、C57BL/6-TREM2- /-和 5XFAD-TREM2 -/- 小鼠大脑中DIMs和DAM的存在和它们的个体发育。应用之前的门控策略(图 4B)并添加针对 CX3CR1、CD11c、CD206 和 CD11a的抗体,作者观察到AD小鼠脑巨噬细胞DIMs(CD83 和 F4/80 高表达,深红点)和DAM(CD11c 高表达和 CX3CR1 低表达,深蓝点)的百分比增加(图 4I 和 4J)。如Keren-Shaul 数据集所述,TREM2- /- 和 AD-TREM2- /-小鼠的 DAM 比例显著降低,而作者发现这两个模型中 DIM比例显著增加(图 4I 和 4J)。大多数DIMs表达大量的Tomato报告基因,证实了它们的单核细胞起源;而 DAM则没有,证实了它们的胚胎起源(图 4K 和 4L)。在表型上与小胶质细胞相比,AD 模型中DIMs的CD45、F4/80、CD83 和 Ly6C 的表达增加,而 DAM 的CD45、CD83、CD206、CD11c 的表达量增加,CX3CR1表达减少(图 4M)。这些观察结果表明,DAM 和 DIMs 是两种在本体发育上不同细胞谱系,可以通过 F4/80、CD83、CX3CR1、CD11c 和 CD206 的表达量来区分,并表现出对TREM2的不同依赖,但两者均在神经退行性变过程中积累。
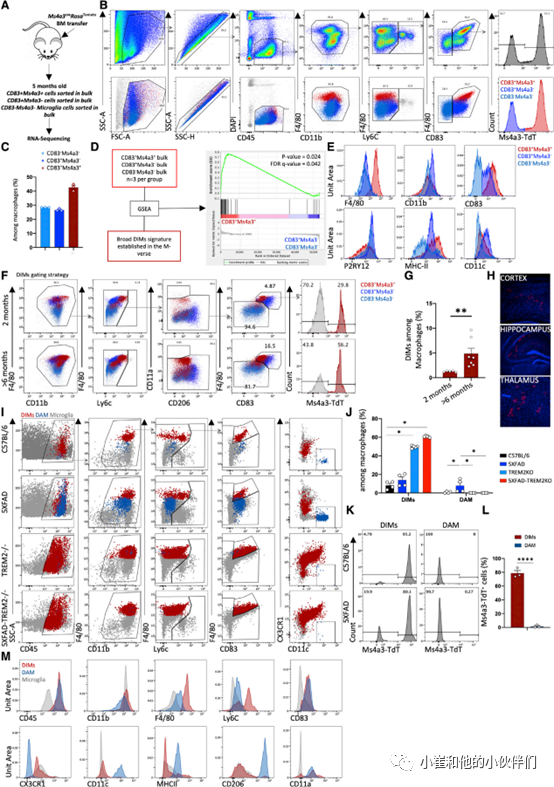
图 4 DIMs来源于单核细胞,而DAM来源于胚胎
9.DIMs与淀粉样β蛋白聚合体在AD小鼠脑内共定位
作者团队接下来使用 5XFAD-Ms4a3CreRosaTomato模型来研究AD大脑中 DIMs的分布情况。通过免疫荧光显微镜研究全脑切片观察到DIMs不是均匀分布的,而是以簇状分布在海马、脑膜、杏仁基底外侧核和皮质内(图 5A 和 5B)。作者还在体内证实了它们的表型:与 Ms4a3-TdT-小胶质细胞相比,Ms4a3-TdT+DIMs表达的 Iba1量相似(图 5C 和 5D),而F4/80 的量显著增加(图 5C 和 5E)。然后,作者团队使用空间密度估计法更详细检查了DIMs 集群,发现 DIMs集群与一些淀粉样蛋白 β (Aβ) 聚集密度高的大脑区域之间的显著重叠(图 5F 和 5G)。在单个细胞水平上,3D 图像重建显示 DIMs与各种大小的 Aβ 聚集体紧密接触,有时几乎与 DIMs本身一样大(图 5H 和 5I)。然而,DIM与Aβ聚集体共定位的程度似乎存在种群水平的限制:局部DIM密度增加,直到达到一定的区域聚集体密度(斑块密度从 0.0010 到 0.0013)。超过该密度DIMs 的数量急剧下降,而 Ms4a3-TdT-小胶质细胞不断聚集(图 5J)。这些结果表明,DIMs 在特定的大脑区域(海马体和基底外侧杏仁核)中积累,与DAM显示在大脑皮层聚集区域不一致。因此,可以在Aβ斑块附近发现DIMs,但只能达到一定的聚集密度,高于此密度的小胶质细胞更可能与Aβ斑块接触。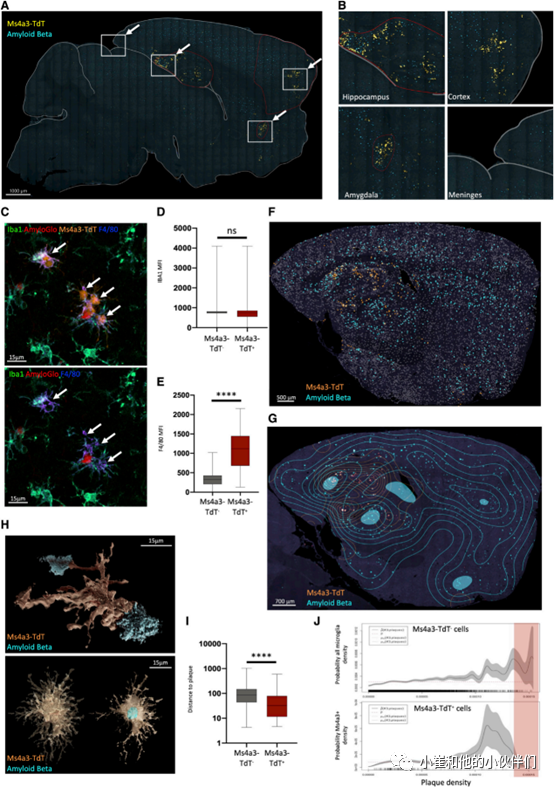
图 5 12月龄5XFAD小鼠特定大脑区域中的DIM聚集
10.DIM、DAM和YAM的等价物存在于人类胎儿和AD大脑中
为了了解这些观察结果与人类大脑的关系,作者团队首先使用Seurat V3重新分析了Thrupp 等人的成人scRNA-seq 脑数据集并确定了18个细胞集群,包括BAMs(MRC1、F13A1)、CD14+和CD16+单核细胞(FCGR1A、FCGR3A、HMGB1)、小胶质细胞(CX3CR1、P2RY12)和污染星形胶质细胞(SNAP25)及少突胶质细胞(PLP1)。当作者更详细地研究小胶质细胞集群时,出现了九个子集群(图 6A),在小鼠 DIMs或 DAM 特征中发现了类似的 DEGs:集群11Up、11Down和9差异表达一些DAM特征基因,而集群12、14和15差异表达一些DIM特征基因(图 6B)。然而,只有集群11Up细胞的CX3CR1和P2RY12的表达显著降低,而GPNMB、SPP1、APOE、TREM2、CD63和ITGAX的表达增加,这与鼠DAM表达谱相对应(图 6B)。此外,与其他集群相比, CD83、EGR1、IER2、FOS、JUN和AXL在12集群中的表达均高于其他集群,对应于鼠DIM表达谱(图 6B)。总体而言,集群11Up 高度表达了鼠广泛DAM特征的20个基因中的8个(B2M、CD63、MAMDC2、CCL3、GPNMB、SPP1、TYROBP 和 TREM2),而集群12 表达了具有广泛 小鼠DIM特征的40个基因中的15个(CCL4、CD14、CD83、CSF2RA、EIF1、FOS、IER2、JUN、JUNB、IL1B、TNF、PLAUR、SAT1 和 BTG2)(图 6C)。然后,作者团队使用集群11Up 和12 的总DEGs来确定这些细胞富集的途径,并再次发现大量重叠:集群11up中前15条途径中的8条也在小鼠DAM中富集(以蓝色突出显示的途径图 6D),集群12中前19条路径中的7条与人类 DIMs共享(图 6D 中以红色突出显示的路径)。 因此,作者认为人类集群11up 可能与鼠DAM相当,而集群12可能与鼠DIMs对应。
为了识别正常发育的大脑中存在的人类YAM、DAM样细胞群,作者团队将 Thrupp数据集与来自Kracht等人的人类胚胎scRNA-seq数据集(13 至 18 周胎儿)整合在一起,并使用与Thrupp数据集相似的参数对Kracht 数据集进行聚类。通过应用上面生成的人类 DIMs(集群 12)和 DAM(集群 11Up)参数,作者不仅能够在集成的Thrupp数据集(图 6E)中识别这些集群,而且还能够在集成的Kracht 数据集中评估它们的相似性(图 6F)。这揭示了来自 Kracht 数据集的胎儿大脑中清晰的DAM样细胞集群,从而代表了人类等效的鼠YAM。并通过查看该集群中细胞的转录数据能够定义人类 YAM 特征(图 6F)。作者团队还在胚胎和老年人的DIMs、DAM和YAM的相对丰度中发现了类似的模式:在成人Thrupp数据集中,DIMs比例较高,DAM 比例较低(图 6G);在胎儿 Kracht数据集中, YAM的比例很高,而DIMs的比例很低(图 6H)。人类YAM 还表达了与鼠YAM相似的代谢途径相关的大量基因,包括与蛋白水解、线粒体组织、谷胱甘肽氧化还原和氧化磷酸化 (OXPHOS) 相关的途径。与鼠类细胞相比,这些细胞进一步显示出更高的脂质改变和降解途径富集。在小鼠和人类YAM普遍富集的细胞过程和信号通路中,作者发现了神经退行性疾病和大脑发育通路,进一步表明 DAM 程序和功能确实是胚胎发育阶段与YAM共享的程序。总之,在小鼠中鉴定的 DIMs、DAM 和 YAM 的scRNA-seq 谱可以扩展到人类,并且这三个不同群体在大脑发育、衰老和神经退行性变过程的程序似乎在这两个物种之间是保守的。
11.CD83+TNF-α+DIMs在AD患者软脑膜中的积累
作者接下来研究DIMs是否表现出人类与AD小鼠模型相同的分布模式和Aβ斑块共定位。通过对AD患者和非AD患者的脑切片进行了 P2RY12、CD83、Aβ和 TNF-α免疫组化,发现AD患者的软脑膜(图6I和6J)中有P2RY12+CD83+细胞,而实质中没有。当观察Aβ和 DIMs的定位时,作者发现Aβ聚集在软脑膜血管周围(图 6J)和脑实质皮层内;同样与非 AD 患者相比,AD 患者的软脑膜中 P2RY12+CD83+TNF-α+细胞的密度明显更高(图 6J 和 6K)。并且与非AD对照相比,AD患者的软脑膜样本中的P2RY12信号更高(图 6K)。此外,与非AD脑相比,AD的软脑膜中表达 DIMs 特征相关分子 AXL 的P2RY12+CD83+细胞密度明显更高(图 6L 和 6M)。因此,表达DIM标志物(包括CD83 和TNF-α)的细胞在AD患者的软脑膜周围积聚,它们可能与AD期间这些区域常见的神经炎症有关。
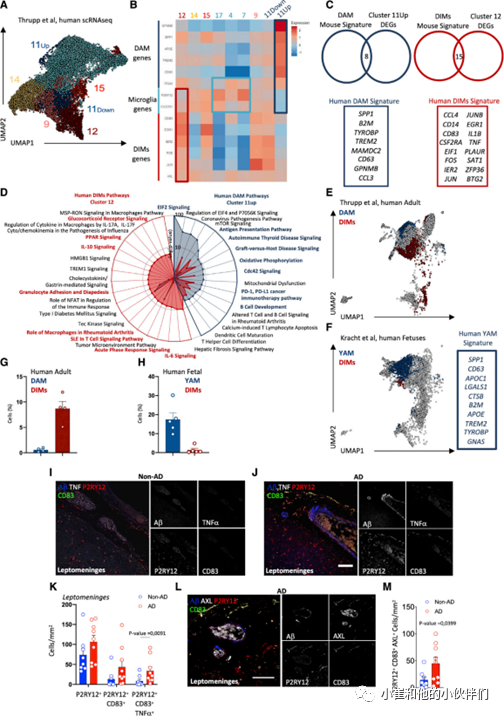
图 6 AD 患者皮质软脑膜中P2RY12+CD83+TNF+细胞数量增加
小结:
通过6个scRNA-seq大脑数据集的整合,作者团队生成了一个横跨胚胎发育到衰老和神经发育的称为 M-Verse的髓系图谱来描绘巨噬细胞群体的异质性。M-Verse 揭示了两种不同的巨噬细胞群,它们表达了已发表的疾病相关小胶质细胞 (DAM) 特征:胚胎衍生的TREM2 依赖性DAM和单核细胞衍生的TREM2非依赖性疾病炎症巨噬细胞 (DIMs)。因此,TREM2作为AD的潜在治疗靶点需要非常谨慎地处理,因为任何变化都可能以目前不可预测的方式影响神经保护性DAM和潜在的神经退行性DIM人群。
: , 。 视频 小程序 赞 ,轻点两下取消赞 在看 ,轻点两下取消在看

